Seo, Kang, Kim, Kwak, Jo, Choi, Boo, and Yoon: Novel rearrangements in the mitochondrial genomes of the Ceramiales (Rhodophyta) and evolutionary implications
ABSTRACT
The Ceramiales is the most diverse and species-rich group (2,669 spp.) of red algae, and it is widely distributed from tropical to polar oceans. Mitochondrial genomes (mitogenomes) and other genes have contributed to our knowledge regarding the classification and phylogeny of this diverse red algal group; however, the mitogenome architecture remains understudied. Here, we compared 42 mitogenomes, including 19 newly generated in this study, to expand our knowledge. The number of genes in mitogenome varied from 43 to 68 due to gene duplication. The mitogenome architecture was also variable, categorized into four types (A–D): type A = ancestral type with a basic composition; type B = those with inverse transpositions; type C = those with inverted duplications; and type D = those with both inversion and duplication. The palindromic and inverted repeats were consistently found in flanking regions of the rearrangement, especially near the cob and nad6 genes. The three rearranged mitogenome architectures (types B, C, D) are the first report of these in red algae. Phylogenetic analyses of 23 protein-coding genes supported the current familial classification of the Ceramiales, implying that the diversity of mitogenome architecture preceded the phylogenetic relationships. Our study suggests that palindromic and inverted repeats may drive mitogenome architectural variation.
Key words: genome rearrangement; mitogenome architecture; molecular evolution; palindromic repeat; red algae
INTRODUCTION
The order Ceramiales is the largest and most diverse group of red algae representing 2,669 species as listed in a global database ( Guiry and Guiry 2023), and this makes the group interesting with regard to its evolutionary history and diversification. The Ceramiales is generally circumscribed by the uniaxial structure of thallus, the presence of periaxial cells, a non-motile male spermatium, a female reproductive structure with a post-fertilization process by the auxiliary cell formation directly from the supporting cell and a triphasic life history ( Kylin 1956, Hommersand 1963, Huisman 2018). Molecular studies have greatly expanded our knowledge of the diversity and phylogeny of the Ceramiales ( Lin et al. 2001, Choi et al. 2002, 2008, Díaz-Tapia et al. 2017, Barros-Barreto et al. 2023). For many years, the Ceramiales employed a four-family scheme: Ceramiaceae, Dasyaceae, Delesseriaceae, and Rhodomelaceae ( Hommersand 1963, Maggs and Hommersand 1993, Womersley 1998). Recently, using plastid genome data from 80 representative species, Díaz-Tapia et al. (2019) proposed the five-family system: Callithamniaceae, Ceramiaceae, Delesseriaceae (including Dasyaceae), Rhodomelaceae, and Wrangeliaceae (segregated from the early diverging Ceramiaceae). The mitochondrial genome (hereafter mitogenome) of red algae includes fast evolving genes in a circular, maternally inherited molecule ( Yang et al. 2015, Salomaki and Lane 2017). A previous study reported the rapid radiation and surprisingly high conservation of mitochondrial gene synteny among the morphologically divergent multicellular lineages of the Rhodymeniophycidae, a subclass of red algae including the Ceramiales ( Yang et al. 2015). Mitogenomes and individual mitochondrial genes have greatly resolved phylogenetic relationships and provided insights into the diversification and its evolutionary histories of red algae (e.g., Hughey et al. 2014, Boo et al. 2016, 2020, Salomaki and Lane 2017, Iha et al. 2018). Previous studies have investigated mitogenome architecture in other groups of cyanidiophycean and florideophycidean red algae ( Lee et al. 2015, 2018, Yang et al. 2015, Cho et al. 2020). In this study, we generated 19 mitogenomes from representative taxa of the order Ceramiales. Specifically, we completed nine mitogenomes for the Ceramiaceae, two each in the Delesseriaceae and Wrangeliaceae, and six in the Rhodomelaceae; however, we were not able to add any taxa from the Callithamniaceae. We focused on the variation of mitogenome architecture from 42 representatives including 23 published mitogenome data in the Ceramiales. From this study, we found that there are four types of mitogenome structure that highlight our knowledge of evolutionary dynamics of the Ceramiales as well as red algae.
MATERIALS AND METHODS
Sample preparation and DNA extraction
Seventeen culture strains of the Ceramiales were obtained from the National Marine Biodiversity Institute of Korea (MABIK). The strains were maintained in the laboratory using L1 enriched seawater medium at 20°C with a 14 : 10 light / dark cycle. Culture tissue (~30 mg) was frozen in liquid nitrogen and ground using Automill TK-AM5 (Tokken Inc., Chiba, Japan). Total genomic DNA was extracted with a modified cetyltrimethylammonium bromide (CTAB) protocol ( Doyle and Doyle 1987). The preheated (65°C) CTAB mixture (CTAB 750 μL, 1% 2-mercaptoethanol 8 μL, and 20 mg mL −1 proteinase K 50 μL) was added to ground tissue and incubated for 10 min at 65°C, followed by a 5 min incubation at room temperature with RNase A (1% v/v). Phenol : chloroform : isoamyl alcohol (25 : 24 : 1) was added in volume equal to sample and centrifuged for 10 min at 14,000 rpm. The supernatant was transferred to a new tube, mixed with 600 μL of chloroform and centrifuged under the same condition. DNA precipitation was achieved by adding 500–600 μL of pre-cooled isopropanol and a volume of 3 M sodium acetate equal to 1/10 of the suspension, followed by incubation at –20°C for 30 min. The supernatant was decanted after centrifugation at 14,000 rpm for 20 min at 4°C. The pellet was washed twice with 1 mL of 70% ethanol, each time inverted and centrifuged at 14,000 rpm for 15 min at 4°C. The final pellet was air-dried and suspended in 50–100 μL of D.W. The quantity and quality of DNA were assessed using the NanoDrop (NABI micro digital, Seongnam, Korea) and Qubit 2.0 fluorometer (Thermo Scientific, Waltham, MA, USA). The concentration of DNA varied in the range of 8–180.8 ng μL −1. The 260/280 absorbance ratios were within the optimal range of 1.9 to 2.1. However, the 260/230 absorbance ratios showed a wider range of 0.97–2.32.
Whole genome sequencing, assembly, and annotation
Whole genome sequencing was conducted using Illumina NovaSeq 6000 (Illumina, San Diego, CA, USA) by DNA-link Inc. (Seoul, Korea). The pair-end sequencing library was prepared using the TruSeq Nano DNA Prep Kit (Illumina), which generated 10 Gb for each taxon. We obtained additional previously generated Illumina read data for Chondria armata (Kützing) Okamura (SRR15927350) and Digenea simplex (Wulfen) C.Agardh (SRR8325636), which were retrieved from the NCBI Sequence Read Archive (SRA). The adapter and low-quality reads were removed using Trimmomatic v0.39 ( Bolger et al. 2014) with the following parameters: ILLUMINACLIP: TruSeq3-PE.fa: 2: 30: 10:2:True LEADING: 3 TRAILING: 3 MINLEN: 36. The mitogenome was assembled using GetOrganelle v1.7.7.0 ( Jin et al. 2020) and NOVOPlasty v.4.3.1 ( Dierckxsens et al. 2017). Genome assemblies obtained with GetOrganelle were manually inspected using Bandage ( Wick et al. 2015). The Illumina reads were mapped to the assembled mitogenomes using Bowtie2 ( Langmead and Salzberg 2012) for validation and error correction. Primary annotation was performed automatically using MFannot ( Beck and Lang 2010). Transfer RNA (tRNA) genes were verified using tRNAscan-SE v.2.0 ( Lowe and Chan 2016). Other possible open reading frames in the intergenic region were searched by using BLASTx. All the predicted genes were confirmed manually by using Geneious Prime 2020.2.4 ( https://www.geneious.com) and BLASTp (e-value, 1e-05). Inverted repeats were identified using Palindrome analyzer ( Brázda et al. 2016). Palindrome analyzer describes the features of palindromic and inverted repeats through similarity, localization, and visualization. Long repeat sequences were analyzed using the Repeat Finder in Geneious Prime with minimum length of 20 bp and 5% of mismatch. The mitochondrial genome sequences have been deposited at GenBank under the accession numbers OR885323–OR885339, BK064963, and BK064964.
Phylogenetic analysis
We collected a total of 42 mitogenomes, including 19 newly generated in this study and 23 from previously published genomes ( Table 1). Rhodymenia pseudopalmata (J.V.Lamouroux) P.C.Silva (KC875852) and Schimmelmannia schousboei (J.Agardh) J.Agardh (KJ398162) were selected as outgroup. Nucleotide sequences of 23 protein-coding genes (PCGs) were aligned with MACSE v2 ( Ranwez et al. 2018) and manually adjusted. The ambiguous region was trimmed with GBlocks v.0.19b ( Castresana 2000) under a 50% gap threshold. Maximum likelihood analysis was performed using IQ-Tree2 v2.0.6 ( Minh et al. 2020) with 1,000 ultrafast bootstrap replications under the GTR + F + I + R5 model. Phylogenetic trees were edited and visualized using FigTree v.1.4.2 ( http://tree.bio.ed.ac.uk/software/figtree).
RESULTS
General feature of mitochondrial genomes
Nineteen newly assembled mitogenomes had an average coverage between 500× and 1,000×. We observed a general decrease in coverage, often to 300×, in palindromic regions. However, the lowest coverage in these regions was >100×, maintaining the integrity and reliability of our data ( Supplementary Figs S1 & S2). Genome sizes and gene contents varied at the family and species level, ranging from 24,508 bp in Campylaephora sungminbooi (J.R.Hughey & G.H.Boo) Barros-Barreto & Maggs (KU145004) to 36,710 bp in Pseudoceramium tenerrimum (G.Martens) Barros-Barreto & Maggs (MABIK-LAS-310); the GC content was 25.0 ± 2.4% ( Table 2). Overall, the mitogenomes contained 43–68 genes, consisting of 21–36 PCGs, 18–33 tRNAs, and 2–4 rRNA subunits ( Table 2). A core set of 23 PCGs were shared in all families, however, up to 36 PCGs were found in the family Ceramiaceae ( Table 2, Fig. 1). Most of the Ceramiales species contained two copies of the ribosomal RNA operon, while, depending on the number of copies of the 5S rRNA, three to four rRNAs were found in five species: Antithamnionella miharai (Tokida) Itono (4 rRNAs), Dasysiphonia japonica (Yendo) S.-H.Kim (3 rRNAs), Bostrychia moritziana (Sonder ex Kützing) J.Agardh (3 rRNAs), Pterosiphonia complanata (Clemente) Falkenberg (3 rRNAs), and Symphyocladiella dendroidea (Montagne) Bustamante, B.Y.Won, S.C.Lindstrom & T.O.Cho (3 rRNAs). All 19 newly analyzed mitogenomes had a group II intron between the nad5 and nad4 genes, and Pseudoceramium tenerrimum contain two introns ( Table 2).
Mitogenome phylogeny
Phylogenomic analysis resolved the relationships of families in the Ceramiales using a concatenated dataset of 23 PCGs (16,263 bp in alignment), excluding duplicated PCGs ( Fig. 2A). The Ceramiales was monophyletic, including four families, which were fully supported. The Rhodomelaceae and Delesseriaceae formed a monophyletic clade, which was a sister to the Wrangeliaceae. The Ceramiaceae diverged first within the Celamiales, which was far related from the remaining families.
Genome rearrangement and expansion in the Ceramiales
We categorized four types of mitogenome architecture in the Ceramiales ( Figs 2B & 3). Type A consisted of 23 PCGs, with 10 PCGs in the forward direction and 13 PCGs of the atp6- nad6 region in reverse direction between the cob and tatC genes. Type A was found throughout the four families and the outgroup species, and here we refer to this as the putative ancestral type. Type B mitogenomes were characterized by having inverse transpositions ( Fig. 3A), as found in Dasya binghamiae A.J.K.Millar and Dasysiphonia japonica. The inverse transpositions occurred in three sets of paired-genes: rps3- rpl16, sdh2- sdh3, tatC- rps12. A deletion of sdh3 gene was found in Dasya binghamiae only. Type C mitogenomes had inverted duplications by repeats, where each pair of duplicated segments revealed a palindromic structure ( Fig. 3B). The duplication was centered on either the 3′ end of the cob or nad6 genes, and the number of duplicated genes varied among species. Type C occurred in four species of the tribe Ceramieae: Campylaephora kondoi (Yendo) Barros-Barreto & Maggs, Campylaeophora sungminbooi, Ceramothamnion sp. MABIK-LAS-337, and Pseudoceramium tenerrimum. The duplication began with the cob gene in Ca. kondoi ( cob- cox3) and Ca. sungminbooi ( cob- rps3). Pseudoceramium tenerrimum and Ceramothamnion sp. MABIK-LAS-337 displayed inverted duplications that started from cob and nad6 genes, respectively. The nad6- nad3 region was duplicated in Ceramothamnion sp. A similar but longer duplication observed in Pseudoceramium tenerrimum, which spanned from nad6 to atp6. Type D mitogenome had both an inversion and a duplication ( Fig. 3B), and this type was found in the tribe Antithamnieae ( Antithamnionella miharai). Specifically, there was an inversion of the genomic segment from nad6 to rps12 genes and duplications occurred for the rrn5- rps11 and nad1 genes.
DISCUSSION
Our mitogenome phylogeny of the Ceramiales is congruent with previous plastid genome-based as well as multigene-based phylogenetic relationships ( Díaz-Tapia et al. 2019, Barros-Barreto et al. 2023). Within the Rhodomelaceae, which is the most species-rich family in the Ceramiales (about 1,100 spp.) ( Guiry and Guiry 2023), the present study confirmed the relationships of 10 tribes. The topology was similar with those of plastid genomes ( Díaz-Tapia et al. 2017), except three were non-monophyletic. For example, Melanothamnus gigas Huisman did not clustered with the other species of the Streblocladieae, Erythrocystis saccata (J.Agardh) P.C.Silva and Vertebrata lanosa (Linnaeus) T.A.Christensen. Gredgaria maugeana Womersley was distantly related to Herposiphonia versicolor (Hooker & Harvey) Reinbold (the tribe Herposiphonieae), but nested in the tribe Pterosiphonieae, making the Pterosiphonieae paraphyletic. This is likely due to low taxon sampling or misidentification. These tribes have been suggested for taxonomic revision in a recent study ( Díaz-Tapia et al. 2023). In the Ceramiaceae, the topology of the tribe Ceramieae is consistent with a recent study based on three-gene phylogeny and morphological characters ( Barros-Barreto et al. 2023). Three new genera, Pseudoceramium Barros-Barreto & Maggs, Yoneshiguea Barros-Barreto, Maggs & M.A.Jaramillo, and Stirkia Barros-Barreto & Maggs were established, and previously synonymized genera, Celeceras Kützing and Reinboldiella De Toni, were reinstated ( Barros-Barreto et al. 2023). However, due to the nomenclatural priority, the genus Ceramothamnion H.Richards was reinstated from the superfluous Stirkia ( Wynne and Schneider 2023). Our comparative analyses of mitogenomes in the Ceramiales demonstrate conserved gene content and synteny, with the exception of the Ceramiaceae. A core set of 23 PCGs was shared in all ceramialean species, with additional PCGs (2–13 genes) resulted from gene duplication in the Ceramiaceae. Two previously published mitogenomes in the Ceramiaceae ( Campylaephora sungminbooi KU145004 and Ceramothamnion japonicum KJ398159) were excluded from the comparative analysis because they were only partially sequenced ( Yang et al. 2015, Hughey and Boo 2016). Hughey and Boo (2016) found a long inverted repeat (about 700 bp) in Ca. sungminbooi, which might disrupt the complete mitogenome assembly of Ca. sungminbooi. The main finding of this study is the identification of four syntenic types of mitogenome in the Ceramiales, of which three of them are newly reported structures in florideophycean red algae. Ancestral type (type A) was widely distributed, including in all four families of the order and outgroup species, Acrosymphytales and Rhodymeniales. The overall gene content and gene synteny of this ancestral type are highly conserved in other reported florideophycidean red algal mitogenomes ( Yang et al. 2015, Lee et al. 2018). However, rpl20 gene has been lost in the Ceramiales as well as in other red algal species, but this lineage-specific gene loss does not correspond to phylogenetic relationships ( Yang et al. 2015). Because three new structures were located within the ancestral type A at different phylogenetic positions (i.e., type B in the Delesseriaceae, type C and type D in the Ceramiaceae), it is highly likely that these three new structures originated independently. It is interesting that these mitogenomic rearrangement typically occurs near the cob and nad6 genes, where palindromic and inverted repeats were found in flanking regions of the rearrangement. The consistent presence of palindromic and inverted repeats in the rearrangement regions suggests that they may initiate or promote these genomic rearrangements. This hypothesis is supported by the observation that palindromic repeats, known to form stable hairpin structures, may cause replication errors and genomic instability ( Cunningham et al. 2003, Miklenić and Svetec 2021). Additionally, the presence of repeated sequences can facilitate genomic rearrangements through unequal crossing over or replication slippage ( Witte et al. 2001, Achaz et al. 2003, Carvalho et al. 2011, Reams and Roth 2015). Therefore, palindromic and inverted repeats of the Ceramiales may initiate the rearrangement of mitogenome structure. However, a question arises: why do other lineages (e.g., the Rhodomelaceae and Wrangeliaceae) not show these syntenic rearrangement, given that ancestral type A still contains palindromic and inverted repeats? We are unable to figure out this mystery; however, a possible scenario is proposed that DNA replication, repair, and recombination-related genes (e.g., RAD52, MSH1) could be involved in unique mitogenome reshaping (e.g., minicircular chromosomes) ( Lee et al. 2023). Nuclear genome analysis would provide some insight into this abnormal phenomenon in future studies. In conclusion, our taxon-wide comparative study of the Ceramiales mitogenomes provides valuable insights into genome architecture with phylogenetic relationships, revealing significant genomic rearrangements and expansions. The palindromic and inverted repeats likely play an important role in genomic rearrangements. The various mitogenome structures of the Ceramiales are first reported in red algae, enhancing our understanding of the diversity and complexity of red algal mitogenome evolution. Our study encourages reexamination of mitogenome architecture in florideophycidean red algae as well as the Callithamniaceae, not covered in the present study, to understand its mechanism and how many variant types occur during the evolutionary process of red algae.
ACKNOWLEDGEMENTS
We thank to Robert A. Andersen for reading and providing valuable corrections in the first version of the manuscript. This work was supported by grants from the National Research Foundation of Korea (grant number NRF-2021R1I1A1A01049542, 2022R1A2B5B03002312, 20-22R1A5A1031361), the Cooperative Research Program for Agriculture Science and Technology Development (Project No. RS-2023-00231243), Rural Development Administration, Republic of Korea, and National Marine Biodiversity Institute of Korea Program (2023M00200).
CONFLICTS OF INTEREST
The authors declare that they have no potential conflicts of interest.
SUPPLEMENTARY MATERIALS
Supplementary Fig. S1
Mapping coverage of established Ceramiales mitogenomes. (A) Antithamnionella miharai (MABIK-LAS-332). (B) Antithamnionella miharai (MABIK-LAS-335). (C) Bostrichia moritziana (MABIK-LAS-101). (D) Bostrychia moritziana (MABIK-LAS-133). (E) Bostrychia tenuissima (MABIK-LAS-110). (F) Campylaephora kondoi (MABIK-LAS-346). (G) Campylaeophora sungminbooi (MABIK-LAS-321). (H) Centroceras gasparrinii (MABIK-LAS-294). (I) Ceramothamnion sp. (MABIK-LAS-337). (J) Chondria armata (SRR15927350) ( https://www.e-algae.org).
Supplementary Fig. S2
Mapping coverage of established Ceramiales mitogenomes. (A) Dasysiphonia japonica (MABIK-LAS-328). (B) Digenea simplex (SRR8325636). (C) Griffithsia okiensis (MABIK-LAS-368). (D) Heterosiphonia pulchra (MABIK-LAS-388). (E) Pleonosporium sp. (MABIK-LAS-307). (F) Polysiphonia morrowii (MABIK-LAS-329). (G) Pseudoceramium inkyui (MABIK-LAS-301). (H) Pseudoceramium tenerrimum (MABIK-LAS-309). (I) Pseudoceramium tenerrimum (MABIK-LAS-310) ( https://www.e-algae.org).
Fig. 1
Presence and absence of mitochondrial protein-coding genes and ribosomal RNAs in the Ceramiales. The order of species follows the phylogeny in Fig. 2. Grey: present, black: 2 copies, white: absent. 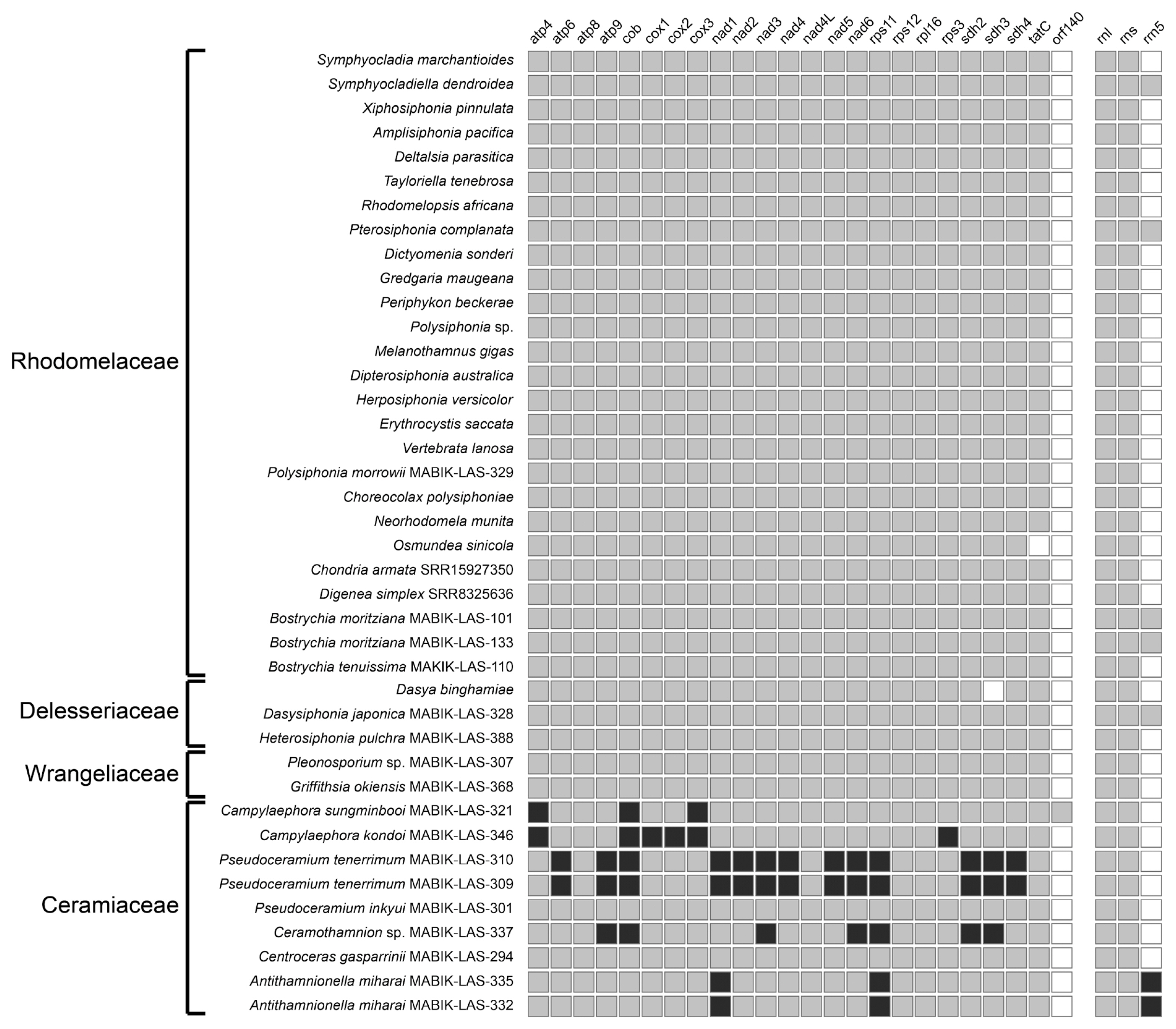
Fig. 2
(A) Maximum likelihood phylogeny of the Ceramiales based on nucleotide sequences of 23 protein-coding genes. Asterisk (*) indicates fully supported bootstrap value. Bold letter indicates newly generated sequences in this study. Deles, Delesseriaceae; Wrang, Wrangeliaceae. (B) Synteny of mitochondrial genome. Four types of mitochondrial structure and gene classification are colored as shown in the key. 
Fig. 3
Syntenic differences among mitochondrial genome in the order Ceramiales. (A) Type B (inverse transpositions) was found in the family Delesseriaceae. (B) Type C (inverted duplications) and type D (inversion and duplications) were found in the family Ceramiaceae. Type A is a putative ancestral type. 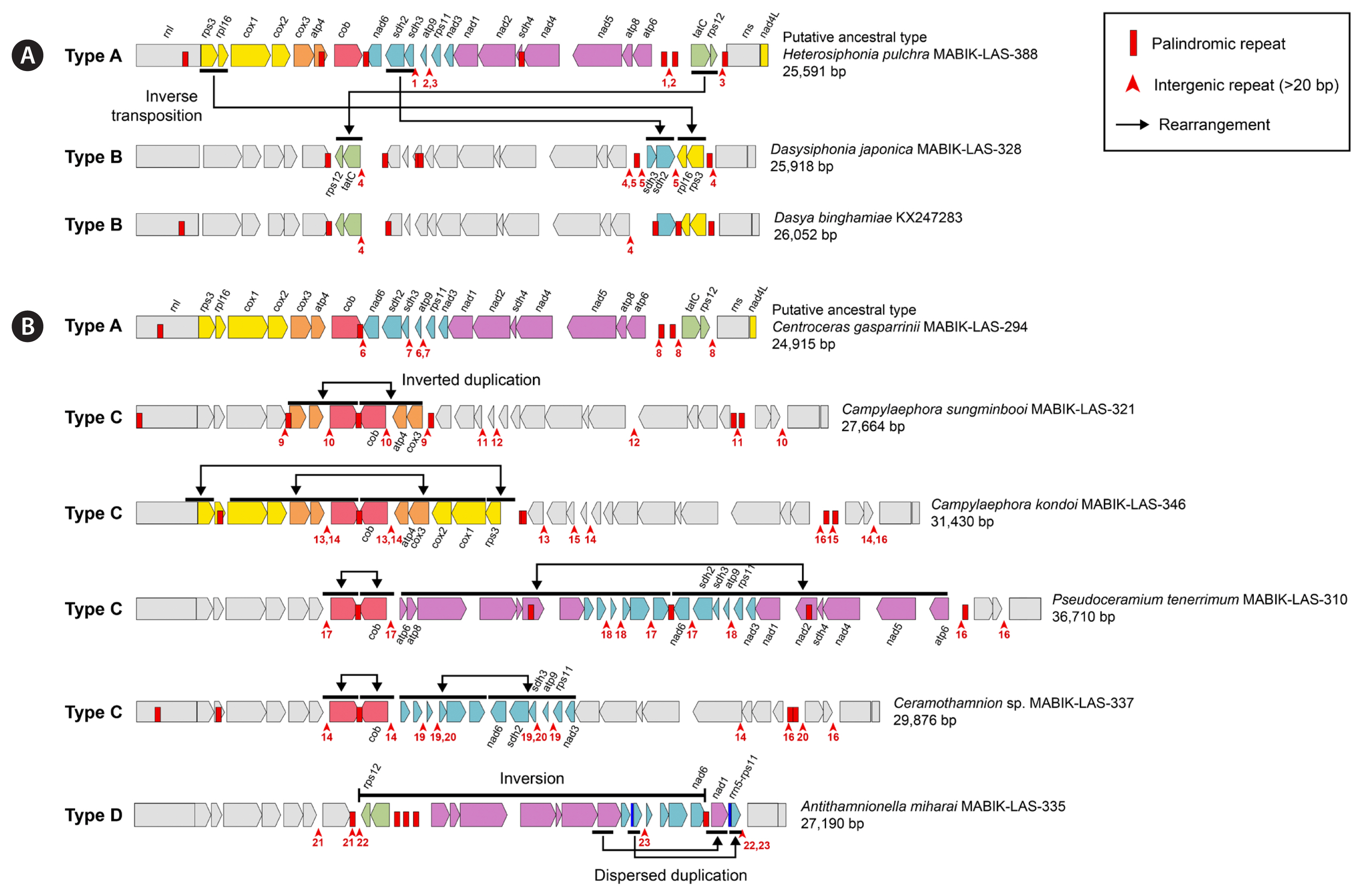
Table 1
Sample list used in phylogenomic analyses including published mitochondrial genomes
|
Family |
Tribe |
Species |
Culture code |
Collection site; latitude, longitude |
GenBank accession No. |
Reference |
|
Ceramiaceae |
Antithamnieae |
Antithamnionella miharai
|
MABIK-LAS-332 |
Daecheon beach, Boryeong, Korea; 36°17′57.69″ N, 126°31′00.33″ E |
OR885323
|
In this study |
|
Antithamnionella miharai
|
MABIK-LAS-335 |
Daecheonhang, Boryeong, Korea; 36°19′30.46″ N, 126°29′58.88″ E |
OR885324
|
In this study |
|
Ceramieae |
Campylaephora kondoi
|
MABIK-LAS-346 |
Euihang-Beach, Taean, Korea; 36°50′33.39″ N, 126°10′19.72″ E |
OR885328
|
In this study |
|
Campylaephora sungminbooi
|
MABIK-LAS-321 |
Jangpyeongri, Tongyeong, Korea; 34°52′20.14″ N, 128°28′02.80″ E |
OR885336
|
In this study |
|
Campylaephora sungminbooi
|
- |
Egerslev Røn, Limfjorden, Denmark |
KU145004 |
Hughey and Boo (2016)
|
|
Centroceras gasparrinii
|
MABIK-LAS-294 |
Guryongpo Beach, Pohang, Korea; 35°59′42.21″ N, 129°33′59.79″ E |
OR885329
|
In this study |
|
Ceramothamnion japonicum
|
- |
Gijang, Busan, Korea |
KJ398159 |
Yang et al. (2015)
|
|
Ceramothamnion sp. |
MABIK-LAS-337 |
Dumunpo, Dolsando(east), Yeosu, Korea; 34°38′43.51″ N, 127°47′50.55″ E |
OR885337
|
In this study |
|
Pseudoceramium inkyui
|
MABIK-LAS-301 |
Janggilri, Pohang, Korea; 35°56′59.02″ N, 129°32′33.82″ E |
OR885335
|
In this study |
|
Pseudoceramium tenerrimum
|
MABIK-LAS-309 |
Myeongsasipri, Wando, Korea; 34°19′31.20″ N, 126°48′15.60″ E |
OR885338
|
In this study |
|
Pseudoceramium tenerrimum
|
MABIK-LAS-310 |
Myeongsasipri, Wando, Korea; 34°19′31.20″ N, 126°48′15.60″ E |
OR885339
|
In this study |
|
Delesseriaceae |
- |
Dasya binghamiae
|
- |
Haida Gwaii, British Columbia, Canada |
KX247283 |
Tamayo and Hughey (2016)
|
|
- |
Dasysiphonia japonica
|
MABIK-LAS-328 |
Songjeong, Gijang, Busan, Korea; 35°10′32.30″ N, 129°11′52.08″ E |
OR885330
|
In this study |
|
- |
Heterosiphonia pulchra
|
MABIK-LAS-388 |
Woohak-ri, Geumodo, Yeosu, Korea; 34°30.43′ N, 127°46.30′ E |
OR885332
|
In this study |
|
Rhodomelaceae |
- |
Amplisiphonia pacifica
|
- |
Haida Gwaii, British Columbia, Canada |
OP748284 |
Díaz-Tapia et al. (2023)
|
|
- |
Deltalsia parasitica
|
- |
Skomer Island, Wales, UK |
OP748283 |
Díaz-Tapia et al. (2023)
|
|
- |
Rhodomelopsis africana
|
- |
Shelley Beach, South Africa |
OP748274 |
Díaz-Tapia et al. (2023)
|
|
- |
Tayloriella tenebrosa
|
- |
Hermanaus, South Africa |
OP748271 |
Díaz-Tapia et al. (2023)
|
|
Alsidieae |
Digenea simplex
|
SRR8325636 |
Onna, Okinawa Prefecture, Japan; 26°29′39.90″ N, 127°50′22.90″ E |
BK064964
|
In this study |
|
Bostrychieae |
Bostrychia moritziana
|
MABIK-LAS-101 |
Tooradin, Western Port Bay, VIC, Australia; 38°14′02.10″ S, 145°24′43.30″ E |
OR885325
|
In this study |
|
Bostrychieae |
Bostrychia moritziana
|
MABIK-LAS-133 |
Tempusak (near Kota Belud), Sabah, Malaysia; 6°23′12.35″ N, 16°20′49.71″ E |
OR885326
|
In this study |
|
Bostrychieae |
Bostrychia tenuissima
|
MAKIK-LAS-110 |
Broughton Ck., NSW, Australia; 34°46′30.12″ S, 150°49′06.22″ E |
OR885327
|
In this study |
|
Chondrieae |
Chondria armata
|
SRR15927350 |
Kyushu Island, Japan; 31°06′36.00″ N, 130°18′04.09″ E |
BK064963
|
In this study |
|
Dipterosiphonieae |
Dipterosiphonia australica
|
- |
Killornei, Victoria, Australia |
OP748281 |
Díaz-Tapia et al. (2023)
|
|
Herposiphonieae |
Gredgaria maugeana
|
- |
The Rip, Victoria, Australia |
OP748280 |
Díaz-Tapia et al. (2023)
|
|
Herposiphonieae |
Herposiphonia versicolor
|
- |
Sant Leonards, Victoria, Australia |
OP748279 |
Díaz-Tapia et al. (2023)
|
|
Laurencieae |
Osmundea sinicola
|
- |
Eureka, near La Paz, Mexico |
MH898940 |
Hughey and Miller (2021)
|
|
Polysiphonieae |
Polysiphonia morrowii
|
MABIK-LAS-329 |
Tappori, Geoje, Korea; 34°45′50.56″ N, 128°35′59.69″ E |
OR885334
|
In this study |
|
Polysiphonieae |
Polysiphonia sp. |
- |
Coral Bay, Western Australia, Australia |
OP748276 |
Díaz-Tapia et al. (2023)
|
|
Pterosiphonieae |
Dictyomenia sonderi
|
- |
Green Head, Western Australia, Australia |
OP748282 |
Díaz-Tapia et al. (2023)
|
|
Pterosiphonieae |
Periphykon beckerae
|
- |
Barrow Island, Western Australia, Australia |
OP748277 |
Díaz-Tapia et al. (2023)
|
|
Pterosiphonieae |
Pterosiphonia complanata
|
- |
Ártabra, A Coruña, Spain |
OP748275 |
Díaz-Tapia et al. (2023)
|
|
Pterosiphonieae |
Symphyocladia marchantioides
|
- |
Praia do Populo, Azores, Portugal |
OP748273 |
Díaz-Tapia et al. (2023)
|
|
Pterosiphonieae |
Symphyocladiella dendroidea
|
- |
- |
OP748272 |
Díaz-Tapia et al. (2023)
|
|
Pterosiphonieae |
Xiphosiphonia pinnulata
|
- |
Bastiagueiro, A Coruña, Spain |
OP748270 |
Díaz-Tapia et al. (2023)
|
|
Rhodomeleae |
Choreocolax polysiphoniae
|
- |
Beavertail State Park, Jamestown, Rhode Island, USA |
KX687877 |
Salomaki and Lane (2017)
|
|
Rhodomeleae |
Neorhodomela munita
|
- |
Badaguan Area, Qingdao, Shandong Province, China |
MW750196 |
Jiang et al. (2021)
|
|
Streblocladieae |
Erythrocystis saccata
|
- |
Pebble Beach, California, USA |
MW810348 |
Amos et al. (2021)
|
|
Streblocladieae |
Melanothamnus gigas
|
- |
Barrow Island, Western Australia, Australia |
OP748278 |
Díaz-Tapia et al. (2023)
|
|
Streblocladieae |
Vertebrata lanosa
|
- |
Beavertail State Park, Jamestown, Rhode Island, USA |
KX687880 |
Salomaki and Lane (2017)
|
|
Wrangeliaceae |
Griffithsieae |
Griffithsia okiensis
|
MABIK-LAS-368 |
Namae Beach, Yangyang, Korea; 37°56′46.81″ N, 128°47′12.73″ E |
OR885331
|
In this study |
|
Spongoclonieae |
Pleonosporium sp. |
MABIK-LAS-307 |
Myeongsasipri, Wando, Korea; 34°19′31.20″ N, 126°48′15.60″ E |
OR885333
|
In this study |
Table 2
General features of the mitochondrial genomes used in the present study
|
Family |
Species |
Size (bp) |
No. of total genes |
Protein-coding gene |
rRNAs |
tRNAs |
Group II intron |
G + C (%) |
|
Ceramiaceae |
Antithamnionella miharai MABIK-LAS-332 |
26,796 |
55 |
25 |
4 |
26 |
1 |
28.3 |
|
Antithamnionella miharai MABIK-LAS-335 |
27,190 |
58 |
25 |
4 |
29 |
1 |
28.3 |
|
Campylaephora kondoi MABIK-LAS-346 |
31,430 |
57 |
29 |
2 |
26 |
1 |
30.2 |
|
Campylaephora sungminbooi MABIK-LAS-321 |
27,664 |
55 |
27 |
2 |
26 |
1 |
29.3 |
|
Centroceras gasparrinii MABIK-LAS-294 |
24,915 |
48 |
23 |
2 |
23 |
1 |
27.6 |
|
Ceramothamnion sp. MABIK-LAS-337 |
29,876 |
65 |
30 |
2 |
33 |
1 |
26.9 |
|
Pseudoceramium inkyui MABIK-LAS-301 |
25,516 |
48 |
23 |
2 |
23 |
1 |
28.6 |
|
Pseudoceramium tenerrimum MABIK-LAS-309 |
36,678 |
68 |
36 |
2 |
30 |
2 |
28.5 |
|
Pseudoceramium tenerrimum MABIK-LAS-310 |
36,710 |
68 |
36 |
2 |
30 |
2 |
28.5 |
|
Delesseriaceae |
Dasysiphonia japonica MABIK-LAS-328 |
25,918 |
49 |
23 |
3 |
23 |
1 |
23 |
|
Heterosiphonia pulchra MABIK-LAS-388 |
25,591 |
47 |
23 |
2 |
22 |
1 |
22 |
|
Rhodomelaceae |
Bostrychia moritziana MABIK-LAS-101 |
25,508 |
45 |
23 |
3 |
19 |
1 |
25.6 |
|
Bostrychia moritziana MABIK-LAS-133 |
25,401 |
45 |
23 |
3 |
19 |
1 |
25.6 |
|
Bostrychia tenuissima MAKIK-LAS-110 |
25,423 |
45 |
23 |
2 |
20 |
1 |
27.9 |
|
Chondria armata SRR15927350 |
25,853 |
49 |
23 |
2 |
24 |
1 |
22.6 |
|
Digenea simplex SRR8325636 |
25,141 |
48 |
23 |
2 |
23 |
1 |
24 |
|
Polysiphonia morrowii MABIK-LAS-329 |
25,235 |
47 |
23 |
2 |
22 |
1 |
23.9 |
|
Wrangeliaceae |
Griffithsia okiensis MABIK-LAS-368 |
24,865 |
46 |
23 |
2 |
21 |
1 |
22.1 |
|
Pleonosporium sp. MABIK-LAS-307 |
24,963 |
48 |
23 |
2 |
23 |
1 |
25.8 |
REFERENCES
Amos, D., Aguilar, V., Barber-Scott, K., Bustamante, D. E., Calderon, M. S., Carrasco, R., Carrion, J. V., Castro, N., Celso, D., Cedillo, S. M. C., Cortes, R., Dao, L., De Santos, S., Ebie, Z., Evangelista, L., Fernandez, S. L., Flores, G., Garcia, L., Gonzalez, E., Hernandz, A., Hernandez, M. O., Hughey, J. R., Luna, L., Marquez, K., Martinez, V., Mendoza, J. E., Mirassou, L., Murillo, C., Parr, M. Jr, Perez, J., Perez-Santana, I., Perez, H., Quezada, A., Quizon, S., Sandberg, S., Santos, A., Tapia, J., Tineo, D. & Vang, M. N. 2021. Transfer of the marine red alga Erythrocystis saccata (Rhodomelaceae, Rhodophyta) to the tribe Streblocladieae inferred from organellar genome analysis. Phytotaxa. 507:266–270.   Barros-Barreto, M. B., Jaramillo, M. A., Hommersand, M. H., Ferreira, P. C. G. & Maggs, C. A. 2023. Phylogenetic analysis of the red algal tribe Ceramieae reveals multiple morphological homoplasies but defines new genera. Cryptogam. Algol. 44:13–58. Boo, G. H., Zubia, M., Hughey, J. R., Sherwood, A. R., Fujii, M. T., Boo, S. M. & Miller, K. A. 2020. Complete mitochondrial genomes reveal population-revel patterns in the widespread red alga Gelidiella fanii (Gelidiales, Rhodophyta). Front. Mar. Sci. 7:583957 pp.
Brázda, V., Kolomazník, J., Lýsek, J., Hároníková, L., Coufal, J. & Št’astný, J. 2016. Palindrome analyser: a new web-based server for predicting and evaluating inverted repeats in nucleotide sequences. Biochem. Biophys. Res. Commun. 478:1739–1745.  Carvalho, C. M B., Ramocki, M. B., Pehlivan, D., Franco, L. M., Gonzaga-Jauregui, C., Fang, P., McCall, A., Pivnick, E. K., Hines-Dowell, S., Seaver, L. H., Friehling, L., Lee, S., Smith, R., Del Gaudio, D., Withers, M., Liu, P., Cheung, S. W., Belmont, J. W., Zoghbi, H. Y., Hastings, P. J. & Lupski, J. R. 2011. Inverted genomic segments and complex triplication rearrangements are mediated by inverted repeats in the human genome. Nat. Genet. 43:1074–1081.  Castresana, J. 2000. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 17:540–552. Cho, C. H., Park, S. I., Ciniglia, C., Yang, E. C., Graf, L., Bhattacharya, D. & Yoon, H. S. 2020. Potential causes and consequences of rapid mitochondrial genome evolution in thermoacidophilic Galdieria (Rhodophyta). BMC Evol. Biol. 20:112 pp. Choi, H.-G., Kraft, G. T., Kim, H.-S., Guiry, M. D. & Saunders, G. W. 2008. Phylogenetic relationships among lineages of the Ceramiaceae (Ceramiaes, Rhodophyta) based on nuclear small subunit rDNA sequence data. J. Phycol. 44:1033–1048. Choi, H.-G., Kraft, G., Lee, I. K. & Saunders, G. 2002. Phylogenetic analyses of anatomical and nuclear SSU rDNA sequence data indicate that the Dasyaceae and Delesseriaceae (Ceramiales, Rhodophyta) are polyphyletic. Eur. J. Phycol. 37:551–569. Díaz-Tapia, P., Maggs, C. A., West, J. A. & Verbruggen, H. 2017. Analysis of chloroplast genomes and supermatrix inform reclassification of the Rhodomelaceae (Rhodophyta). J. Phycol. 53:920–937. Díaz-Tapia, P., Pasella, M. M., Verbruggen, H. & Maggs, C. A. 2019. Morphological evolution and classification of the red algal order Ceramiales inferred using plastid phylogenomics. Mol. Phylogenet. Evol. 137:76–85. Díaz-Tapia, P., Rodríguez-Buján, I., Maggs, C. A. & Verbruggen, H. 2023. Phylogenomic analysis of pseudocryptic diversity reveals the new genus Deltalsia (Rhodomelaceae, Rhodophyta). J. Phycol. 59:264–276. Dierckxsens, N., Mardulyn, P. & Smits, G. 2017. NOVOPlasty: de novo assembly of organelle genomes from whole genome data. Nucleic Acids Res. 45:e18 pp. Doyle, J. J. & Doyle, J. L. 1987. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 19:11–15.
Guiry, M. D. & Guiry, G. M. 2023. Algae Base. World-wide electronic publication, National University of Ireland, Galway, Available from: http://www.algaebase.org
. Accessed Sep 15, 2023 Hommersand, M. H. 1963. The morphology and classification of some Ceramiaceae and Rhodomelaceae. Univ. Calif. Publ. Bot. 35:165–366.
Hughey, J. R. & Boo, G. H. 2016. Genomic and phylogenetic analysis of Ceramium cimbricum (Ceramiales, Rhodophyta) from the Atlantic and Pacific oceans supports the naming of a new invasive Pacific entity Ceramium sungminbooi sp. nov. Bot. Mar. 59:211–222. Hughey, J. R., Gabrielson, P. W., Rohmer, L., Tortolani, J., Silva, M., Miller, K. A., Young, J. D., Martell, C. & Ruediger, E. 2014. Minimally destructive sampling of type specimens of Pyropia (Bangiales, Rhodophyta) recovers complete plastid and mitochondrial genomes. Sci. Rep. 4:5113 pp. Hughey, J. R. & Miller, K. A. 2021. Genetic investigation of three type specimens of Osmundea (Rhodomelaceae, Rhodophyta) from the Gulf of California, Mexico and California, USA. Phytotaxa. 489:65–78. Huisman, J. 2018. Algae of Australia. 2. Red algae. ABRS & CISRO Publishing, Canberra, 672 pp.
Iha, C., Grassa, C. J., Lyra, G. M., Davis, C. C., Verbruggen, H. & Oliveira, M. C. 2018. Organellar genomes: a useful tool to study evolutionary relationships and molecular evolution and Gracilariaceae (Rhodophyta). J. Phycol. 54:775–787. Jiang, Z., Li, R., Cui, Y., Jia, X., Liu, T., Wang, X. & Qu, J. 2021. The complete mitochondrial genome and phylogenetic analysis of Neorhodomela munita
. Mitochondrial DNA B Resour. 6:2746–2747. Jin, J.-J., Yu, W.-B., Yang, J.-B., Song, Y., dePamphilis, C. W., Yi, T.-S. & Li, D.-Z. 2020. GetOrganelle: a fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 21:241 pp. Kylin, H. 1956. Die Gattungen der Rhodophyceen. C.W.K. Gleerups, Lund, 673 pp.
Lee, J.-M., Boo, S. M., Mansilla, A. & Yoon, H. S. 2015. Unique repeat and plasmid sequences in the mitochondrial genome of Gracilaria chilensis (Gracilariales, Rhodophyta). Phycologia. 54:20–23. Lee, J. M., Song, H. J., Park, S. I., Lee, Y. M., Jeong, S. Y., Cho, T. O., Kim, J. H., Choi, H.-G., Choi, C. G., Nelson, W. A., Fredericq, S., Bhattacharya, D. & Yoon, H. S. 2018. Mitochondrial and plastid genomes from coralline algae provide insights into the incongruent evolutionary histories of organelles. Genome Biol. Evol. 10:2961–2972. Lee, Y., Cho, C. H., Noh, C., Yang, J. H., Park, S. I., Lee, Y. M., West, J. A., Bhattacharya, D., Jo, K. & Yoon, S. H. 2023. Origin of minicircular mitochondrial genomes in red algae. Nat. Commun. 14:3363 pp. Lin, S.-M., Fredericq, S. & Hommersand, M. H. 2001. Systematics of the Delesseriaceae (Ceramiales, Rhodophyta) based on large subunit rDNA and rbc L sequences, including the Phycodryoideae, subfam. nov. J. Phycol. 37:881–899. Maggs, C. A. & Hommersand, M. H. 1993. Seaweeds of the British Isles. 1. Rhodophyta. Part 3A. Ceramiales. Natural History Museum, London, 444 pp.
Minh, B. Q., Schmidt, H. A., Chernomor, O., Schrempf, D., Woodhams, M. D., von Haeseler, A. & Lanfear, R. 2020. IQ-TREE 2: new models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 37:1530–1534. Salomaki, E. D. & Lane, C. E. 2017. Red algal mitochondrial genomes are more complete than previously reported. Genome Biol. Evol. 9:48–63. Tamayo, D. A. & Hughey, J. R. 2016. Organellar genome analysis of the marine red alga Dasya binghamiae (Dasyaceae, Rhodophyta) reveals an uncharacteristic florideophyte mitogenome structure. Mitochondrial DNA Part B Resour. 1:510–511. Womersley, H. B. S. 1998. The marine benthic flora of southern Australia - Part IIIC. Ceramiales
- Ceramiaceae, Dasyaceae
. Australian Biological Resources Study & State Herbarium of South Australia, Canberra & Adelaide, , 535 pp.
Wynne, M. J. & Schneider, C. W. 2023. Reinstatement of Ceramothamnion H. Richards (1901), a replacement name for the newly described Stirkia (Ceramiaceae, Rhodophyta). Not Algarum. 296:1–4.
Yang, E. C., Kim, K. M., Kim, S. Y., Lee, J., Boo, G. H., Lee, J.-H., Nelson, W. A., Yi, G., Schmidt, W. E., Fredericq, S., Boo, S. M., Bhattacharya, D. & Yoon, H. S. 2015. Highly conserved mitochondrial genomes among multicellular red algae of the Florideophyceae. Genome Biol. Evol. 7:2394–2406.
|
|











