INTRODUCTION
Because of industrialization, the atmospheric carbon dioxide (CO2) concentration increased by about 50%, from ~280 ppm at the beginning of the industrial era to 413 ppm at present (September 2021) (NOAA/GML 2021). This increase in atmospheric CO2 concentration, along with other greenhouse gases, is linearly correlated with the rise of global and oceanic temperatures (NOAA/NCEI 2021). At the same time, a significant buildup of dissolved CO2 in the upper layer of ocean leads to increased bicarbonate (HCO3−) concentration, which brings about a decrease in seawater pH by increasing H+ concentration. Consequently, a perturbation of the carbonate buffering system and a process collectively referred to as ocean acidification (OA) occur (Zeebe et al. 2008, Doney et al. 2009). It has been projected that OA will cause the average pH of the upper pelagic ocean to drop by 0.3–0.4 units, which is equivalent to 150% increase in H+ and 50% decrease in carbonate ion (CO32−) availability (Orr et al. 2005, Gattuso et al. 2015). Also, a drop in the pH in coastal waters by about 0.45 units is predicted by the end of 2100 (Cai et al. 2011). Although atmospheric inputs are not the only carbon source, this can be used to reliably project the drop in pH of coastal waters as about 12% faster than that in epipelagic oceans. The variability of pH, which is driven by complex physical and biological processes, is often substantially higher in the coastal waters as it is relatively more affected by anthropogenic factors such as eutrophication and organic carbon runoff (Carstensen and Duarte 2019, Ding et al. 2019, He and Silliman 2019).
Notable diurnal or spatial variations in the pH of coastal waters of the Korea Strait, important for commercial shellfisheries and aquaculture, can range between 6.6 and 8.2 (Kim et al. 2012). However, this range is unrealistic because the lowest pH value of the Yellow Sea over the past 30 years (1985–2014) was estimated to be 7.6 (Kim et al. 2020). This OA is crucial for marine algae relying on dissolved inorganic carbon (DIC) for photosynthesis and / or calcification and represents a significant threat to marine habitat-forming species, ecosystem function, and associated services (Iglesias-Rodriguez et al. 2008, Hurd et al. 2009, Kroeker et al. 2013, Brondizio et al. 2019, Zunino et al. 2021). The ranges of physiological characteristics of marine organisms can be linked to OA conditions due to altered inorganic carbonate chemistry resulting from increased HCO3− and H+, and decreased CO32− and calcium carbonate saturation states (Doney et al. 2009, Kroeker et al. 2013, Riebesell and Gattuso 2015). The higher availability of DIC resulting from OA might be promoting greater photosynthesis and growth of marine phytoplankton that are less efficient in carbon sequestration. For example, many phytoplankton species are positively affected by OA as their productivity, growth rate, and carbon sequestration increasing significantly (Rost et al. 2008, Beardall et al. 2009, Basu and Mackey 2018, Lim et al. 2020). Conversely, reduced pH and CO32− can pose a substantial challenge for marine calcifiers that build their skeletons from calcium carbonate and for species which are sensitive to higher proton levels during photosynthesis (Guinotte and Fabry 2008, Comeau et al. 2013, Kottmeier et al. 2016).
Most marine phytoplanktons, including dinoflagellates, have evolved with sophisticated carbon concentrating mechanisms (CCMs) that help them acquire sufficient CO2 for photosynthesis. CCM is one of the several types of metabolic processes that facilitate active influx of CO2 and / or bicarbonate ions (HCO3−) to overcome the limited CO2 availability in the vicinity of RuBisCO (ribulose-1,5-bisphosphate carboxylase / oxygenase) (Raven et al. 2017). Photosynthetic dinoflagellates contain form II RuBisCO that converts carbon dioxide to reduced organic compounds with a poor affinity for CO2 (Reinfelder 2011). However, dinoflagellates are not carbon-limited at the current CO2 levels due to effective CCMs (Fu et al. 2008, van de Waal et al. 2019). They could also benefit from OA by downscaling their CCMs and reallocating energy and resources towards other cellular compartments and biological processes (Rost et al. 2008).
Non-calcifying algae are more likely to benefit from OA, although non-CCM species that depend exclusively on CO2 availability are carbon-limited (Cornwall and Hurd 2016, Ji et al. 2017). For example, OA has been shown to enhance the growth rates of dinoflagellates Amphidinium carterae (Bausch et al. 2019), Alexandrium fundyense (Hattenrath-Lehmann et al. 2015), Alexandrium tamarense (Pang et al. 2017), and Karlodinium veneficum (Fu et al. 2012). Growth rates of bacillariophytes Phaeodactylum tricornutum (Laws et al. 1997) and Thalassiosira pseudonana (Sobrino et al. 2008), raphidophytes Chattonella subsalsa (Fu et al. 2008), and Heterosigma akashiwo (Hennon et al. 2017), and cyanobacteria Prochlorococcus (Bagby and Chisholm 2015) grown in elevated pCO2 environments also increased, while it did not affect the photosynthesis and growth of some species with active CCMs (Cornwall and Hurd 2016). The growth of macroalgae with active CCMs, such as Ulva prolifera and Sargassum muticum, could be enhanced by OA (Gao et al. 2016, Xu et al. 2017). In contrast, adverse effects on growth were observed in dinoflagellates Alexandrium monilatum and Prorocentrum minimum (Hennon et al. 2017) and bacillariophyte Phaeodactylum tricornutum (Laws et al. 1997). For calcifying phytoplankton, such as coccolithophores, it is challenging to overcome enhanced dissolution due to lowered saturation state of calcium carbonate associated with the decrease in carbonate ion (CO32−) concentrations caused by OA (Joint et al. 2011, Reusch and Boyd 2013, Hutchins and Fu 2017).
In general, growth rates of phytoplankton increase with warming to a point (optimum temperature) beyond which they decrease sharply. Species from thermally fluctuating environments, like temperate regions, have a broad thermal niche and biogeography, and therefore can tolerate thermal stress or are less sensitive to warming (Pörtner et al. 2017). While many studies have examined the combined effects of OA and warming on phytoplankton ecology (Feng et al. 2009, Kim et al. 2013, Maugendre et al. 2015, Paul et al. 2016, Bénard et al. 2018, Gao et al. 2018), no study has explored the effects of OA on the thermal performance curve (TPC) of growth and chain formation over a wide range of temperatures.
Photosynthetic organisms may acclimate to changed carbonate chemistry within short periods through phenotypic plasticity and genotypic variations for long-term adaptation (Hutchins and Fu 2017, Ding et al. 2019). To project the future trends of proliferation of harmful algal blooms (HAB) species, it is crucial to understand their physiological and morphological traits, which are highly temperature-dependent (Lee et al. 2019b, Kang et al. 2020, Kruk et al. 2021). Although high temperatures may negatively impact phytoplankton production and biomass, marine species can mitigate some ecological impacts of short-term heatwave and warming events through rapid adaptation and phenotypic plasticity in response to regional environments (Listmann et al. 2016, O’Donnell et al. 2018, Kang et al. 2019b, Lee et al. 2020b). However, these responses of HAB species to OA caused by changing carbonate chemistry still lack experimental evidence.
Alexandrium affine has been reported from arctic to tropical waters (Guiry and Guiry 2021). This ubiquitous species is not considered a paralytic shellfish toxin-producing species, but it produces lytic compounds and other allelochemicals (Basti et al. 2015). In the Korea Strait, A. affine appears throughout the year with an increased abundance during spring to summer, immediately after diatom bloom termination (Lim et al. 2019, Lee et al. 2020a). In contrast, A. pacificum which has the sxtA gene is reported to occur in certain locations in the Pacific region, such as China, Japan, Korea, Australia, and New Zealand (Genovesi et al. 2015, Shin et al. 2017, Dai et al. 2020). In the Korea Strait, A. pacificum appears from late January to early March (Lee et al. 2020a). Thus, differences in biogeography and phenology between two Alexandrium species are anticipated.
Many studies have detailed positive, negative, or neutral impact of OA on the physiology, growth, and morphology of dinoflagellates. However, the effects of OA on Alexandrium species are still uncertain. This study aims to compare the effects of elevated pCO2 on photosynthesis, growth, and chain-forming thermal performances of A. affine and A. pacificum, which have caused notable economic losses in the Korean shellfish industry during the last decades.
MATERIALS AND METHODS
Species and culturing
Two species of Alexandrium isolated from the coastal waters of the Korea Strait were used. Alexandrium affine (Aa-MEDHO0710) and A. pacificum (Ap-MEDHO0210) were isolated from Mijo Harbor (34°42′ N, 128°03′ E) in September 2007 and Jangmok Bay (34°59′ N, 128°40′ E) in February 2010, respectively. Cultures of these strains were maintained in f/2 medium without silicate at 20–23°C and 32–34 psu under a 12 : 12 h light-dark cycle with ~100 μmol photons m−2 s−1, typically containing a mixture of single cells and two- and more than three-cell chains. Alexandrium species-specific primer sets targeting the internal transcribed spacer region (5′-TGTGCTTGACTTTTACATGA-3′ and 5′-GAGCAGCACAGATACA-3′ for A. affine and 5′-TTCAATGCAAAACATTGACCT-3′, and 5′-CTGTTTGCATTTCTCTAGTT-3′ for A. pacificum) were applied to verify the identification of species by conventional PCRs (Supplementary Fig. S1).
Experimental setup
Cells of A. affine and A. pacificum were acclimated in batch mode to the experimental temperatures and pCO2 levels for 14 days before the start of the experiment. Subsequently, A. affine and A. pacificum were cultured semi-continuously with the abundance of 100 cells mL−1 and 200 cells mL−1, respectively. Compared with target pH values, these relatively low cell densities allowed us to maintain the pH with minor variations (<0.10 units). Cells incubated in 250-mL Erlenmeyer flasks at 33–34 psu under an incident irradiance of 150 μmol photons m−2 s−1 provided by four 36W daylight fluorescent lamps (Dulux L 36W/86; Osram, Münich, Germany), with a light-dark cycle of 14 : 10 h.
The five target temperatures were controlled using a thermostatically controlled water bath, which allowed control of the temperature within ±0.5°C (Supplementary Fig. S2). The temperature was logged every 30 min using data loggers (Onset Computer Corporation, Pocasset, MA, USA). A given target pCO2 level was manipulated by adding an appropriate amount of CO2-saturated seawater (CSS), no more than 10 μL. CSS was obtained by bubbling reagent-grade CO2 through seawater in gas-tight containers with the pH dropping below 5. Samples were collected in triplicates from each semi-continuous culture for pHT measurements to reset the target pCO2 level. The photosynthetic efficiency and cell counts of the samples were also measured.
All inoculums were transferred weekly to keep them in the exponential growth phase until the start of the experiment. During the semi-continuous cultivation in Erlenmeyer flasks (100 mL capacity), cultures were maintained at the lower section of the exponential phase (Supplementary Fig. S3), and were optically thin to minimize self-shading, CO2 drift, and nutrient limitation.
Inorganic carbonate chemistry
The inorganic carbonate chemistry was subsequently calculated with the help of CO2SYS program (Lewis and Wallace 1998) using the total pH scale (pHT), total alkalinity (AT), temperature, and salinity of culture seawater (Lim et al. 2020). The pHT on the total hydrogen scale was monitored for each culture every day using a high-resolution UV-VIS spectrophotometer (Agilent 8453; Agilent Technologies, Pal Alto, CA, USA) fitted with 20-mm path length special optical glass spectrophotometric cells (18B-SOG-20, Starna). Absorbances of 30 mL seawater samples kept at 25°C were measured after adding 80 μL aliquots of m-cresol purple indicator dye (Sigma-Aldrich Chemical Co., St. Louis, MO, USA) at wavelengths 434, 578, and 730 nm.
Total alkalinity (AT) measurements of each culture were obtained at the beginning and the end of the 10-day incubation using potentiometric titration with acid in a semi-closed cell system, consisting of a Metrohm 765 Dosimat titrator (Metrohm, Zofingen, Switzerland) connected to a pH meter (Orion 920A; Thermo Fisher Scientific, Waltham, MA, USA). The electromotive force during titration with acid (0.13 N HCl) was recorded using Q-basic software (Millero et al. 1993). AT values were calibrated against certified seawater reference material (Andrew Dickson, Scripps Institution of Oceanography).
Growth rate and chain formation
Live cells were enumerated every day in a 1 mL Sedgewick-Rafter counting chamber on an inverted microscope (IX73; Olympus, Tokyo, Japan). Specific growth rates (μ) were calculated for each replicate of the applied treatments by fitting an exponential function through all cell counts over time as follows:
, where Nt denotes the cell count at time t, and N0 denotes the cell count at the start of each experiment.
Lugol-fixed samples from the harvest day were used for the measurement of chain-forming cells using an inverted microscope (IX73; Olympus). For each replicate, at least 100 single or chain cells were randomly assigned for each treatment.
Cardinal temperature estimation
The Cardinal Temperature Model with Inflection (CTMI) in which the maximum specific growth rate (μmax) is described as a function of temperature (T, °C) alone was used (Rosso et al. 1993). The cardinal temperature parameters were obtained by fitting the CTMI defined as follows:
In CTmin ≤ T ≤ CTmax, where CTmin (°C) and CTmax (°C) are the hypothetical lower (minimum) and upper (maximum) critical temperatures, respectively, through which the growth rate is zero. However, we represented CTmin and CTmax as the temperatures at which the growth rate was 5% of the maximum growth rate due to their ambiguity, that determined the temperature tolerance range (Ttol). The thermal performance breadth (TB80) was also defined as the 80th percentile of the CTMI fits where the performance was close to optimal. Topt (°C) is the temperature at μmax, and μopt is the specific growth rate at Topt.
Maximum photochemical efficiency of photosystem II
To determine photosynthetic activity, the maximum efficiency of photosystem II (PSII) photochemistry (i.e., the maximum quantum yield of PSII) was determined using a pulse amplitude-modulated (PAM) chlorophyll-a fluorometry (Phyto-PAM fluorometer; Walz GmbH, Effeltrich, Germany). The maximum quantum yield (Fv/Fm) for each culture was measured after being acclimatized to darkness for 15 min. Where Fv (variable fluorescence) is calculated as the difference between Fo and Fm fluorescence, where Fo is the minimum level of fluorescence in the absence of photosynthetic light at open reaction centers, and Fm is the maximum fluorescence yield induced by a saturation light pulse at closed reaction centers of PSII (Genty et al. 1989).
Statistical analyses
For experimental measures (carbonate chemistry, maximum efficiency of PSII photochemistry, growth rate, and chain formation), the main and interaction effects of temperature (five levels ranging from 15 to 31°C for A. affine or 30°C for A. pacificum) and pCO2 (ambient and elevated levels) to assess significant responses were evaluated by one- or two-way analyses of variance (ANOVA). Data were initially checked for conformity to normality using the Shapiro-Wilk test, and for homogeneity of variances using Levene’s test. If a significant ANOVA was found, differences among the group means were evaluated using Turkey’s post hoc tests for multiple comparisons. All statistical analyses were performed in SPSS Statistics version 25 (IBM Corp., Armonk, NY, USA).
RESULTS
The carbonate chemistry levels between the ambient and elevated pCO2 conditions were significantly different for all temperature levels during experiments (p < 0.05) (Tables 1 & 2, Fig. 1), despite a slight upward drift in pH during active cell growth. For instance, mean pCO2 ranged from 459 to 637 μatm in ambient and 1,030 to 1,082 μatm in elevated pCO2 conditions in A. affine cultures (Table 1). In A. pacificum cultures, the mean pCO2 in ambient and elevated pCO2 ranged from 302 to 373 μatm and 885 to 1,047 μatm, respectively (Table 2). The other dissolved carbonate species showed a similar trend.
Comparison between species in the ambient condition
The values of Fv/Fm (maximum quantum efficiency of PSII) for A. affine gradually increased as temperature increased from 15 to 23°C and remained high at 27 and 31°C. For A. pacificum, the Fv/Fm was highest at 23°C, and showed a clear decreasing trend beyond this temperature (Fig. 2).
The shape of TPCs and cardinal temperature parameters differed between the species. For instance, the maximum growth rate (μmax) of A. affine observed at an optimal temperature (Topt) of 26°C was 0.54 d−1, which is faster than A. pacificum that had an average growth rate of 0.51 d−1 at a Topt of 25.3°C (Fig. 3). Critical thermal minimum (CTmin) and critical thermal maximum (CTmax) were 15.6 and 35.3°C, respectively, for A. affine, and 7.0 and 30.7°C, respectively, for A. pacificum. Thermal breadth (TB80) also differed between the species, with the values 8.4 and 7.8°C for A. affine and A. pacificum, respectively (Table 3).
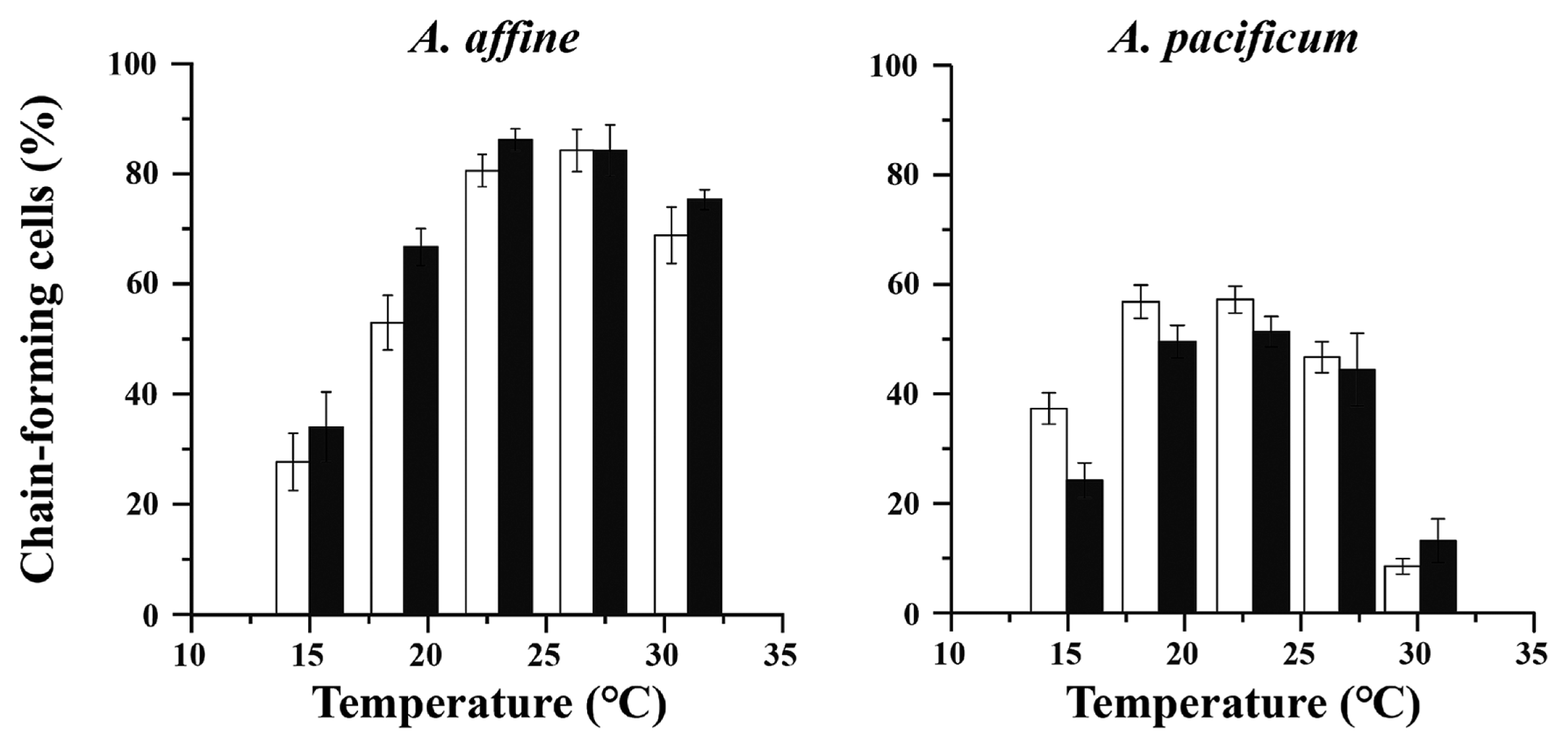
Percentage of chain-forming cells varied from 28 to 84% at 15 and 27°C for A. affine, and from 8 to 57% at 30 and 23°C for A. pacificum (Fig. 4). A. pacificum showed a lower proportion of cells with chains than A. affine. There were also differences in chain-length fractionation (CF) between the species (Fig. 5). A. affine showed more and various CF than A. pacificum, with a range of 5–40% of short (2–3 cells), middle (4–8 cells), and long (>8 cells) chains against 2–35% of primarily short CF across the experimental temperature range.
Comparison between CO2 conditions
There was no significant interaction between temperature and pCO2 on the Fv/Fm for both species. However, A. affine was significantly affected by temperature only (p < 0.001), and A. pacificum was significantly affected by both temperature and pCO2 (p < 0.001) (Fig. 2, Supplementary Table S1). Exposure to elevated pCO2 lead to only minor increases in the Fv/Fm of A. affine within a range of 0.4 to 0.8, with the most prominent negative effect being observed at the lowest temperature (15°C) (Fig. 2). Compared to A. affine, the values of Fv/Fm of A. pacificum were somewhat lower at temperatures beyond the optimal temperatures under both ambient and elevated pCO2 conditions.
A. affine showed a significant increase in growth rate as a response to elevated pCO2 compared to ambient conditions (p < 0.001) (Supplementary Table S2). In the same way, its maximum growth rate (μmax) increased by 11% (from 0.54 to 0.60 d−1), CTmax increased from 35.3 to 37.1°C, and CTmin decreased from 15.6 to 14.9°C, while Topt remained unchanged at 26.0°C for both ambient and elevated pCO2 conditions. Thermal breadth (TB80) increased from 8.4 to 10.2°C only under elevated pCO2 conditions. For A. pacificum there was no significant effect of elevated pCO2 on the growth rate (p = 0.489) (Supplementary Table S2), even its maximum growth rate (μmax) increased from 0.51 d−1 under ambient condition to 0.58 d−1 under elevated pCO2 conditions. Topt decreased from 25.3 to 24.6°C, CTmin increased from 7.0 to 9.8°C, and CTmax decreased from 30.7 to 30.4°C under elevated pCO2 when compared with ambient conditions. Thermal breadth (TB80) also decreased from 7.8 to 7.3°C when such comparison was made. The TB80 indicates that A. affine was not stressed by the experimental treatments, pointing to its broad temperature tolerance resulting in a wide fundamental niche in respect to the future climate.
The chain-forming capacity did not vary significantly between ambient and elevated pCO2 conditions (p > 0.01), but significant differences between temperatures occurred (p < 0.001) (Supplementary Table S3). The percentage of chain-forming cells was higher in A. affine than in A. pacificum, that found to markedly increase at their optimal growth temperatures (Fig. 4). A. affine exposed to elevated pCO2 responded by building up chains of various lengths and had higher (but not significantly higher) proportion of chain-forming cells relative to that of ambient pCO2 cultures. In contrast, A. pacificum had only a few cell chains with a slightly higher proportion in ambient than in elevated pCO2 conditions, showing that chain formation was not related to CO2 treatment.
We also tested whether species responded differently to elevated pCO2 for chain-forming trait, and found that only A. affine exhibited a more substantial increase in the proportion of middle chain-length (4–8 cells chain−1) fractionation, while no change in short chain-length (2–3 cells chain−1) and long chain-length (>8 cells chain−1) fractionation was found under elevated pCO2 conditions (Fig. 5, Supplementary Table S4). In A. pacificum, elevated pCO2 did not cause a change in CF (Fig. 5, Supplementary Table S5).
DISCUSSION
Experimental constraints
The pH drift and CO2 exchange in the culture flask were dependent on the growth rate and concentration of cells. In nature, pH and pCO2 may vary considerably algal blooming, with drastic increases in pH and reductions in CO2 levels at the bloom peak (Brandenburg et al. 2021). It is unclear whether cells become inhibited by high pH or limited by low CO2 during the experiment; nevertheless, by allowing pH drift to occur, the experiment represents changes in carbonate chemistry due to biomass build-up due to some treatments. During the semi-continuous culture, flasks were kept at the lower section of the exponential growth phase (Supplementary Fig. S3) and were maintained at, or below, the starting cell concentration of ca. 100 cells mL−1 for A. affine and ca. 200 cells mL−1 for A. pacificum to avoid self-shading, nutrient limitation, and minimize CO2 exchange. Such experimental cell concentrations are far away from those found in bloom episodes in nature.
Photosynthetic activity
The maximum quantum yield (Fv/Fm) has been used as a stress indicator in phototrophic organisms for characterizing the physiological status (Kim et al. 2019). Two Alexandrium tended to increase their Fv/Fm in response to elevated pCO2, but elevated pCO2 had a significant effect only on the Fv/Fm of A. pacificum (Supplementary Table S1). The Fv/Fm of A. affine showed neither a significant difference between pCO2 conditions nor considerable deterioration across temperature gradients during the experiment. It indicates that A. affine is not stressed by the combined treatments, pointing to tolerate broad temperature resulting in a wide fundamental niche in respect to those environmental factors. Various studies support this and generally describe Alexandrium species as eurythermal marine species (Lee et al. 2019a, Lim et al. 2019). Also, an overall increase of the Fv/Fm as considered a marker of its photosynthetic ability reflects its current health status. However, Chl-a fluorescence analysis is not always a reliable method when considering an identifiable Fv/Fm to measure the potential for photosynthetic activity. Note that these measurements represent only a snapshot of bio-optical characteristics of PSII photochemistry that occur within the reaction center complex of PSII (van de Poll et al. 2020).
Thermal performance for growth rate and cardinal temperatures
There was a clear difference between the species in their growth performance response to elevated pCO2 (Fig. 3). Overall, the growth rates of A. affine showed an apparent increase under elevated pCO2, whereas A. pacificum showed a slight, insignificant increase (Supplementary Table S2). The optimal temperature (Topt) of A. pacificum decreased from 25.3°C under ambient to 24.6°C under elevated pCO2 conditions and was lower than that of A. affine (estimated at 26.0°C under both conditions) (Table 3). The sharp decline in growth rate above the Topt of A. pacificum indicates its high sensitivity to heat stress once Topt is surpassed during summer days in the shallow waters of Korea Strait. In contrast, A. affine had a higher critical thermal maximum (CTmax) compared to A. pacificum, which might indicate a more extraordinary competitive ability of A. pacificum to withstand heatwaves prevalent during warm season in the Korea Strait. This could be why the abundance of A. pacificum in the Korea Strait does not increase in summer (Lim et al. 2019, Lee et al. 2020a). Although A. affine is a seasonal species that blooms during the spring-summer months, it is surprising a low abundance of species remains in deep-water of the Korea Strait during the winter months (Lee et al. 2020a), which could have shaped its wide thermal tolerance range (Ttol) and thermal performance breadth (TB80).
In this laboratory experiment, thermal breadth (TB80) of A. affine widened by 21%, while that of A. pacificum narrowed by 6% with elevation of pCO2 levels (Table 3, Fig. 3). Thus, A. affine can be considered as a thermal generalist species, which can tolerate and grow under a wide range of temperatures, with a wide geographical distribution (Farrell 2009). Since both species share habitat and locality in the Korea Strait, the considerable variation in cardinal temperatures may reflect thermal niche partitioning between species, likely arising from enhanced resource use efficiency and interaction in the temperate pelagic system. Two Alexandrium species increased their maximum growth rate (μmax) in response to elevated pCO2 (Table 3). Based on the observed changes in cardinal temperatures, we anticipate the abundance of A. affine to eventually increase in the Korea Strait, given the warming of surface waters and its synergistic effect with increasing pCO2.
Elevating CO2 concentration would favor RuBisCO’s carboxylation reaction over oxygenation activity, and further gains in photosynthesis would be granted. The role of CCM is to suppress photorespiration and support adequate carbon fixation by enhancing the chemical conditions in the vicinity of the primary CO2 assimilating enzyme RuBisCO (Raven et al. 2017). Although we did not measure CCM activity, dinoflagellates with a high CCM activity also revealed a high CCM plasticity (van de Waal et al. 2019). Therefore, Alexandrium also may possess effective and flexible CCMs. Especially A. affine with high growth rate may have down-regulated their CCMs in the high CO2 condition to reallocate energy towards other cellular processes such as carbon metabolism, cell growth, and chain formation.
Chain formation
Most phytoplankton species maintain a relatively uniform morphology over time, but others actively change their size by forming chains. Chains form when daughter cells do not separate at the end of cell division, generating highly elongated shapes. Cell size and shape of phytoplankton can be mediated by many essential processes, including light capture, nutrient acquisition, predation, and motility (Pahlow et al. 1997, Smayda 2010, Selander et al. 2011, Bergkvist et al. 2012, Bjærke et al. 2015, Lovecchio et al. 2019). The swimming speed of phytoplankton increases with chain length because chain formation increases propulsion force rather than hydrodynamic drag (Fraga et al. 1989). Chain-forming dinoflagellates, such as Alexandrium species, were observed to swim much faster than single cells (Fraga et al. 1989, Selander et al. 2011, Lovecchio et al. 2019) and to have the ability to migrate vertically between surface waters, during the daytime to harvest light, and deep waters, during the night to consume nutrients (Ryan et al. 2010). Chain length plasticity has been suggested to provide adequate refuge to phytoplankton to reduce encounters with grazers and regulate the balance between motility and grazer avoidance (Selander et al. 2011, Bergkvist et al. 2012, Kang et al. 2019a, Jang and Jeong 2020, You et al. 2020). However, our results from laboratory cultures suggest that maximum capacity of chain formation by Alexandrium species is attained in the absence of grazing pressure, which occurs in the absence of chemical cues released by the grazers. Although the chain formation extent of A. affine and A. pacificum showed varied responses with temperature, no significant effects of pCO2 on the chain formation of both species were observed (Fig. 4, Supplementary Table S3). Moreover, chain length correlates to the growth rate in both species, suggesting that chain length might also depend on growth conditions. In addition, an enhanced response of chain length was observed only in A. affine, showing that elevated pCO2 significantly enhances the middle CF (4–8 cells chain−1) (Fig. 5). Differential chain formation responses of A. affine vs. A. pacificum indicate species-specific responses in physiological adjustment and carbon allocation strategies for their growth to cope with OA.
In conclusion, the Alexandrium species are cryptic, sharing similar morphological characteristics, and show phenotypic plasticity within traits in response to elevated pCO2. Two Alexandrium can acclimate their physiology to local environmental conditions within their plastic response range. Although the strains were isolated from the same region, the two species differed strongly in their chain-forming capacity, cardinal temperatures, and response directions under elevated pCO2.