INTRODUCTION
Pyropia yezoensis (Bangiales, Rhodophyta) is a commercially valuable and most cultivated marine red alga. It has a heteromorphic life cycle, alternating between the foliose thallus gametophyte and filamentous sporophyte generations (Blouin et al. 2011). The gametophyte thalli, which are used as an important food resource, grow in cold water during winter and transitions to the sporophytic conchocelis phase in summer. Temperature is one of the major abiotic stresses affecting P. yezoensis gametophyte growth rate (Avila et al. 1986, Hwang et al. 1997, Luo et al. 2014, Hwang and Park 2020). Therefore, the rise in seawater temperature owing to global warming is expected to significantly limit Pyropia growth. To mitigate the effects of increasing seawater temperatures, it may be necessary to develop high-temperature-resistant varieties of Pyropia.
Understanding the molecular genetic circuits of plants exposed to high temperatures will be key to successful breeding of heat-resistant crop varieties. Plant responses to heat stress include a reduction in the expression level of genes involved in photosynthesis, the biosynthesis of storage compounds, such as starch, and an increase in the expression level of the genes that are important for cellular homeostasis (Shinozaki et al. 2015). Heat shock proteins (HSPs) are typically genes that are upregulated under high-temperature conditions and play an important role in the cellular response to heat stress (Wang et al. 2004). HSPs act as molecular chaperones that prevent irreversible aggregations and re-solubilize already aggregated proteins under heat stress conditions (Wang et al. 2004). Heat stress results in the formation of reactive oxygen species (ROS), which at low concentrations play a role as signaling molecules but at high concentrations may lead to oxidative damage. To protect against this damage, the levels of ROS scavengers such as superoxide dismutases, catalases, and peroxidases in the cells are increased (Mittler 2002, Mittler et al. 2004). In addition, compatible solutes, such as proline, glycine betaine, and sugar alcohols, are accumulated in the cells, similar to those observed in the response to osmotic stress (Mittler 2002, Livingston et al. 2009, Van den Ende and Valluru 2009, Dai et al. 2020). Furthermore, previous studies have demonstrated that heat stress affects the structure of the cell wall (Moore et al. 2008, Sasidharan et al. 2011, Le Gall et al. 2015, Chen et al. 2020); the plant cell wall determines the size and shape of the cell through the mechanical control of cell expansion (Chen et al. 2020). Some cell wall-related genes may play a role in the acquisition of thermotolerance (Yang et al. 2006, Ha et al. 2007, Rienth et al. 2013, Le Gall et al. 2015, Chen et al. 2020). However, additional studies are needed to fully understand the role of the plant cell wall in heat tolerance at the physiological, genetic, and biochemical levels. Our knowledge of the heat response in plants is based on green plants including Arabidopsis, but research on red algae including Pyropia is currently limited.
To find the genes involved in the acquisition of heat tolerance in P. yezoensis, we compared transcriptomes from gametophytes of wild-type and heat-tolerant mutants P. yezoensis under control and heat-stress conditions and identified differentially expressed genes (DEGs). We selected a DEG, PyHRG1, which was downregulated in the gametophytes of wild-type P. yezoensis under high-temperature conditions and in the heat-tolerant mutant, and characterize its physiological function. This study would be valuable by providing the gene resources for heat tolerance in P. yezoensis.
MATERIALS AND METHODS
Plant material and stress treatment
P. yezoensis var. Sugwawon 104 (SG104) and Gy500 were obtained from the Fisheries Seed and Breeding Research Institute, Korea. Gy500 is a heat-tolerant mutant developed from P. yezoensis SG104 (Park et al. unpublished data). Heat-tolerant mutant Gy500 was developed by radiation breeding as described by Shin et al (2018). Conchocelis line from selected heat tolerant gametophyte was established after three rounds of the asexual cycle from single monospore to thallus. Gametophyte thalli of Gy500 grew better than those of SG104 under both high-temperature and normal growth conditions. P. yezoensis was cultured in modified Grund medium (McLachlan 1973) in a growth chamber under the following conditions: temperature, 12°C; irradiation, 80 μmol photon m−2 s−1 provided by cool-white fluorescent lamps; photoperiod, 10 : 14 (light : dark). For heat treatment, growth bottles containing P. yezoensis were transferred to a growth chamber at 20°C with the same light intensity and photoperiod. All gametophyte thalli were snap frozen using liquid nitrogen and maintained at −80°C until RNA extraction.
Transcriptome sequencing and identification of DEGs
Transcriptome sequence reads were obtained in triplicate from gametophyte thalli of wild-type (SG104) and mutant (Gy500) P. yezoensis, under control (12°C) and high-temperature (20°C) conditions. Library construction and RNA sequencing was performed using the Illumina Hi-Seq 2500 platform at G&C Bio Company (Daejeon, Korea). Briefly, sequencing reads were evaluated for quality using FastQC (http://www.bioinformatics.babraham.ac.uk/projects/fastqc/) and preprocessed to remove adapter sequences using Cutadapt software (ver. 1.9.1;https://cutadapt.readthedocs.io/en/stable/).Finally, high-quality, clean reads were subjected to de novo assembly using the Trinity (https://github.com/trinityrnaseq/trinityrnaseq) software with the default parameters.
Fragments per kilobase per million reads (FPKM) values were applied to measure gene expression levels. Sequencing reads from each cDNA library were mapped to the de novo assembled contigs using Bowtie2 (http://bowtie-bio.sourceforge.net/bowtie2/index.shtml), and DEGs were detected using edgeR (https://bioconductor.org/packages/release/bioc/html/edgeR.html), with criteria of at least 2-fold ratio changes and q-values < 1.0 × 10−3. The heatmap of the sample correlation matrix was generated across all replicates using the Trinity-bundled Differential Expression analysis script PtR with default parameter. For gene annotation, DEGs were compared to the sequences in the NCBI non-redundant database (updated Jul 30, 2021) and Uniprot/Swissprot (updated Jul 31, 2021) using BLASTX program 2.12.0+ with default parameter. Contigs with an E-value of >1.0E-10 were considered not available.
Identification and characterization of PyHRG1
Among the selected DEGs, the contig Py97124 had no sequence homology to any known gene and was highly downregulated under heat stress conditions and in the heat-tolerant mutant. The downregulated expression pattern of contig Py97124 was validated using quantitative reverse transcription polymerase chain reaction (qRT-PCR). Therefore, we selected Py97124 for further study and named it “Heat Response Gene 1 of P. yezoensis (PyHRG1).”
The cDNA covering the full open reading frame (ORF) of PyHRG1 was amplified from wild-type (SG104) and mutant (Gy500) P. yezoensis and cloned to create a pGEM T-easy vector (Promega, Madison, WI, USA). The putative molecular weight and isoelectric point (pI) of PyHRG1 were predicted using Geneious R8 software (Biomatters Limited, Auckland, New Zealand). To identify PyHRG1 homologs in the Pyropia genome, amino acid sequences deduced from PyHRG1 were used to search the draft-genome sequence of P. yezoensis (Kim et al. 2021). Multiple sequence alignments of amino acid sequences were performed using ClustalX software (http://www.clustal.org/clustal2). A phylogenic analysis was conducted using the neighbor-joining method using CLC viewer 8.0 (CLC Bio, Aartus, Denmark).
Expression and subcellular localization of PyHRG1
Total RNA was obtained from the gametophyte thalli using an RNeasy plant mini kit (Qiagen, Hilden, Germany), following the manufacturer’s instructions. qRT-PCR was performed as described by Choi et al. (2013) using PyHRG1-specific primer sets (5′-GTCAAAACGCCGACCAAGAC-3′and5′-CACGATCCATCGTCCTGCTT-3′). PyUBQ(5′-TTTCCAAGGTGCTCCTCTCCATC-3′and 5′-CGTCTCTTCATAGCGACTGCGGTT-3′) was used as an internal control. All the samples were run in duplicate, and the n-fold differential expression was calculated using the comparative Ct method (2−ΔΔCt) with three replicates. Differences between the samples were compared using t-test in Microsoft Excel software.
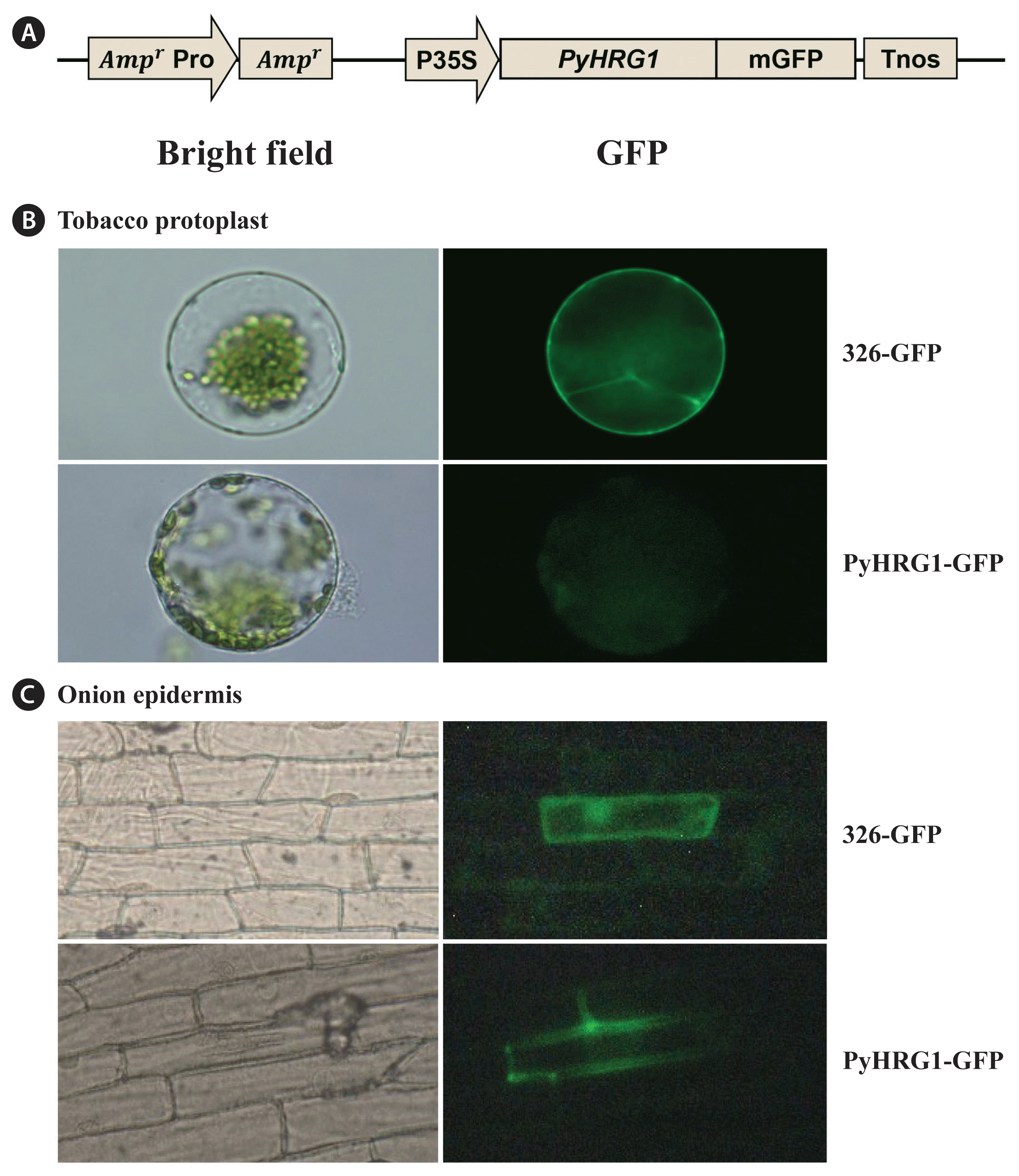
Cellular localization of PyHRG1 was analyzed using the web-accessible software WoLF PSORT (https://wolfpsort.hgc.jp) and DeepLoc (http://www.cbs.dtu.dk). Bioinformatic analysis predicted that PyHRG1 could be a secretory protein (Supplementary Table S1). To examine the cellular localization of PyHRG1, the PyHRG1 coding region was amplified using the primers 5′-TCTAGAATGGTCGGTACCGC-3′ and 5′-GGATCCAGCACTTCTGCCA-3′, containing an XbaI site upstream and a BamHI site downstream. PCR products were introduced into the XbaI and BamHI sites of the 326-GFP-3G vector. The constructed PyHRG1-GFP vector was introduced into tobacco (Nicotina benthamiana) protoplasts. The transformed tobacco protoplasts were then incubated at 25°C for 12–24 h under dark conditions. The recombinant DNA was also introduced into onion (Allium cepa) epidermis cells via particle bombardment, as described by Ha et al. (2007). Subsequently, the fluorescence signals of PyHRG1-GFP fusion protein were evaluated under a fluorescence microscope (Leica, Wetzlar, Germany).
Transformation and abiotic stress tolerance assay of Chlamydomonas
The physiological function of PyHRG1 in the single-cell green alga Chlamydomonas reinhardtii was assayed as described by Im et al. (2017). The ORF of PyHRG1 was amplified using primers 5′-CGCCATATGGTCGGTACCGCCGC-3′ and 5′-CTGCAGGCACTTCTGCCACGATCCA-3′ and then subcloned into the NdeI and PstI sites of the PsaD promoter in pCr112, a Chlamydomonas expression vector. This pCr112-PyHRG1 plasmid was then introduced into C. reinhardtii Mut11. The introduction of PyHRG1 into transgenic Chlamydomonas cells and its expression were confirmed using qRT-PCR. The Chlamydomonas actin gene (CrActin) was used as a control (5′-TGTGCATACGTGGATAGCTTG-3′ and 5′-ATGACCCGCTCCTCATATCTT-3′).
To assay C. reinhardtii cell growth, cells were cultured to an absorbance of 0.75 at 750 nm. C. reinhardtii cells were diluted (10−1 to 10−3) with TAP fresh medium, inoculated onto agar plates, and cultured at 23°C in a growth chamber under a 14 : 10 (light : dark)-photoperiod and cool-white fluorescent light (50 μmol photon m−2 s−1). For the heat treatment, cells were incubated in a growth chamber at 35°C, under the same light intensity and photoperiod. Four days after heat treatment, cells were transferred to a growth chamber kept at 23°C and cultured for 1 week. C. reinhardtii Mut11 harboring an empty pCr112 vector was used as a control.
RESULTS AND DISCUSSION
Transcriptome sequencing and identification of DEGs
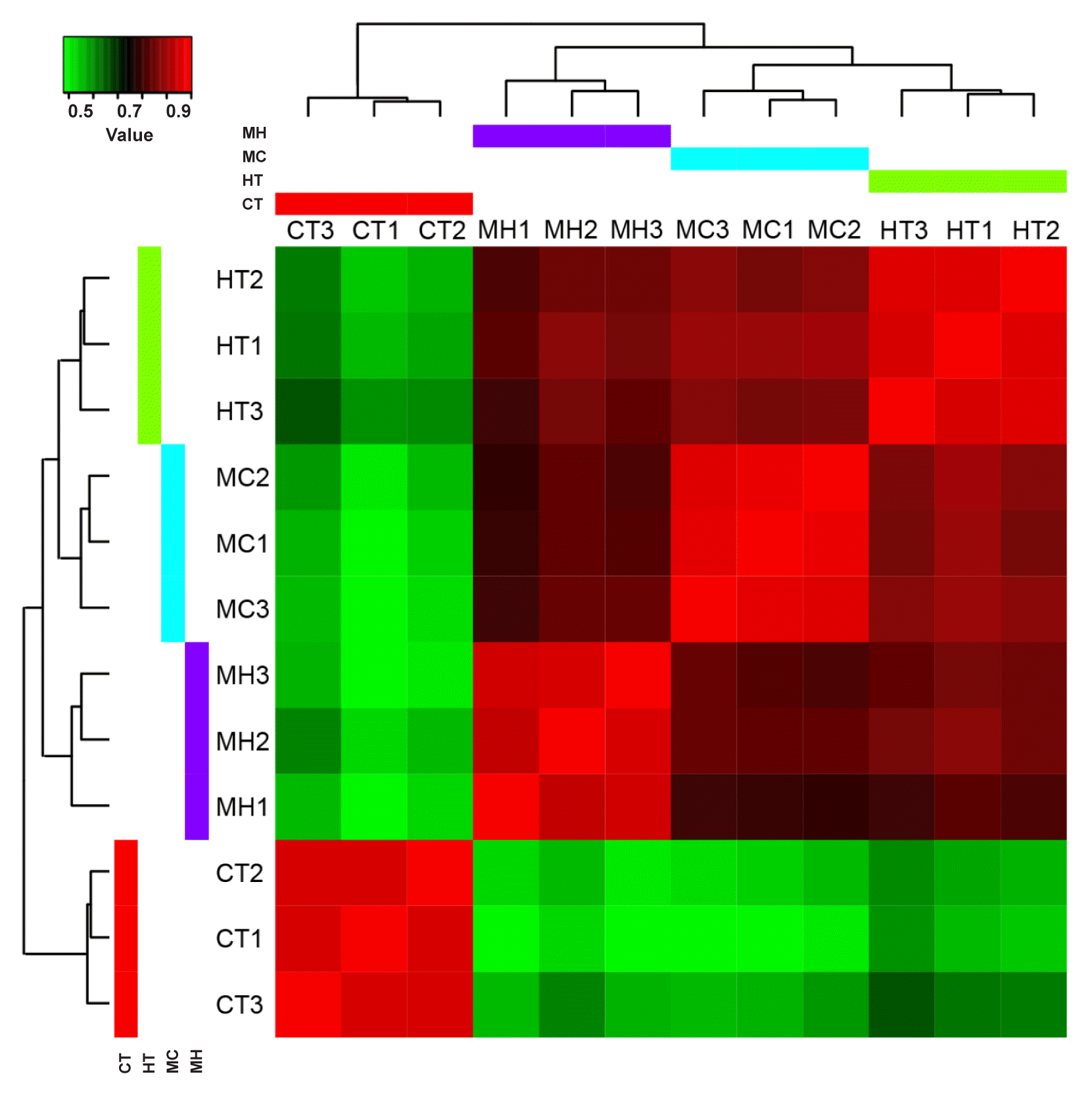
To identify genes that respond to heat stress in P. yezoensis, cDNA libraries were constructed from the gametophytes of wild-type SG104 and heat-tolerant mutant Gy500 under normal culture conditions (control) or heat stress (20°C) conditions. Three cDNA libraries were generated from three replicates for each sample. A total of 113.2 Gb of transcriptome sequences were generated from 12 libraries, and a total of 69.4 Gb of high-quality sequences were obtained after preprocessing (Table 1). De novo assembly processing generated a total of 242,470 contigs, with an N50 length of 961 bp and covering a total of 161.8 Mb (Table 1). Clustering analysis of the sample correlation matrix showed that each replicate was strongly correlated within samples and was reliable (Fig. 1). Transcriptomes of the heat-tolerant mutant Gy500, in both control conditions and under heat stress, were correlated significantly higher with those generated from gametophytes of wild-type SG104 under heat stress than with those generated under control condition (Fig. 1).
To screen for DEGs responsive to heat stress, contigs were analyzed by FPKM value fold change comparisons. A total of 1,251 contigs were identified as DEGs in response to heat stress (Supplementary Tables S1 & S2). A total of 409 contigs among the identified DEGs were downregulated in heat stressed wild-type SG104 and showed low levels even under the control condition in heat-tolerant mutant Gy500 (Supplementary Table S2). Transcriptome analysis is a rapid and efficient method being used to identify genes involved in specific metabolic or stress tolerance processes (Rodriguez et al. 2010, Song et al. 2016, Chen and Li 2017, Wang et al. 2017). RNA sequencing projects have been applied to the phylum Rhodophyta to identify genes involved in the development and abiotic stress responses (Chan et al. 2012, Choi et al. 2013, Im et al. 2015, 2017). Previous studies in Rhodophyta, however, observed transcriptional changes in gametophyte of wild type plants under control and stress condition. Transcriptome sequences from heat-tolerant mutant Gy500 of P. yezoensis will facilitate future studies for identification and understanding of the molecular mechanisms involved in heat stress tolerance in P. yezoensis.
Identification and characterization of PyHRG1
In this study, we focused on DEGs, which were significantly downregulated in the gametophytes of wild-type SG104 under heat stress and also detected at low levels in the heat-tolerant mutant Gy500 under both control and heat stress conditions. DEGs with FPKM values greater than 20 in SG104 under control conditions and SG104 : Gy500 ratios greater than 32 under control conditions were selected and summarized in Table 2. Among them, we selected the DEG Py97124, which was strongly downregulated in the gametophytes of wild-type SG104 under heat stress and detected at low levels in the heat-tolerant mutant Gy500 (Table 2). Downregulation of the DEG Py97124 in gametophyte under heat stress condition and in heat-tolerant mutant were confirmed using qRT-PCR (Supplementary Fig. S1).
The cDNA of PyHRG1 (accession No. MT122996) encodes a polypeptide of 241 amino acids with a molecular weight of 26.5 kDa and a pI of 9.11. Glycine (14.2%) was the most abundant amino acid in the PyHRG1 polypeptide (Fig. 2A). PyHRG1 showed no sequence homology with any known genes currently deposited in public databases, except with those of other red algae; PyHRG1 homologs were also identified from other Pyropia species, P. tenera and P. seriata (Fig. 2A). PyHRG1 shared 97.9% sequence identity with the PtHRG1 of P. tenera, the closest relative of P. yezoensis. PyHRG1 homologs were also identified in another Porphyra species, Porphyra umbilicalis (Fig. 2B). Although red algae and green plants share abiotic stress tolerance mechanisms, not all stress response genes identified in green plants are found in red algae; some stress genes are specific to red algae (Choi et al. 2013, Lu and Xu 2015, Im et al. 2017, Na et al. 2018). Data from this study suggest that PyHRG1 is a novel red algae-specific gene or its homologs in green plants have significantly lower sequence similarity for identification.
Based on the analysis of the draft-genome sequence of P. yezoensis, we identified three PyHRG1 homologs, which shared approximately 31.1–41.2% amino acid sequence identity with PyHRG1 (Supplementary Table S3 & Fig. S2). PyHRG1 homolog 4 had the highest sequence homology with PtDEG5, which was previously reported to be a P. tenera desiccation response gene (Im et al. 2017). These results suggest that PyHRG1 may be involved in responses to desiccation as well as to heat stress. Although its expression patterns were different for SG104 and Gy500, cDNA sequences of PyHRG1 were identical for both P. yezoensis variants (Supplementary Fig. S3). These results suggest that the high-temperature tolerance phenotype of Gy500 is not a result of PyHRG1 mutation but rather a result of downregulation of PyHRG1 expression in Gy500.
PyHRG1 inhibits cell growth in Chlamydomonas reinhardtii
PyHRG1 was downregulated in P. yezoensis SG104 gametophytes under heat stress condition. And in gametophytes of heat-tolerant mutant Gy500, the transcripts of PyHRG1 was detected at a much lower level than that of SG104 (Table 2). To assess the physiological function of PyHRG1, the ORF of PyHRG1 was subcloned into the PsaD promoter, a constitutive expression promoter of Chlamydomonas, in a pCr112 vector. PyHRG1 was introduced into the single-cell green alga, C. reinhardtii Mut11. Introduction and expression of PyHRG1 in transgenic C. reinhardtii were verified using reverse transcription polymerase chain reaction with PyHRG1-specific primers. PyHRG1 transcripts were detected in all selected transformed C. reinhardtii cells, and no amplification bands were observed in control cells transformed with an empty pCR112 vector (Fig. 3A). Transgenic C. reinhardtii cells overexpressing PyHRG1 had slower growth rates than wild-type cells under normal growth condition (23°C) (Fig. 3B). These results demonstrate that PyHRG1 plays a role in the growth of C. reinhardtii cells. When transgenic C. reinhardtii cells overexpressing PyHRG1 were exposed to heat stress, cell growth inhibition was further exacerbated. These results suggest that PyHRG1 is not the only gene involved in cell growth inhibition under heat stress. It is not clear whether PyHRG1 acts directly by inhibiting cell growth or indirectly by affecting other proteins that control cell growth. Studies in green plants have shown that under abiotic stressors, plants alter expression of genes to reduce growth and increase resistance to stress (Wahid et al. 2007, Shinozaki et al. 2015); likewise, in P. yezoensis, PyHRG1 plays a role in cell growth regulation in response to heat stress.
Plants have evolved genetic systems to respond efficiently to adverse heat stress. The duration and severity of stress, susceptibility of cell types, and stage of development all influence the ability of a particular genotype to survive heat stress (Wahid et al. 2007, Shinozaki et al. 2015). HSPs plays critical role in heat tolerance process by prevent irreversible aggregations and re-solubilize proteins that have already aggregated by heat stress (Wang et al. 2004). A wide spectrums of heat response genes were identified by transcriptome analysis (Song et al. 2016, Chen and Li 2017, Wang et al. 2017, Xu and Hwang 2018). Besides HSPs, information of the molecular and physiological function of heat response genes from red algae is limited.
Subcellular localization of PyHRG1
Bioinformatics analysis predicted that PyHRG1 could be a secretory protein (Supplementary Table S4). To examine the cellular localization of PyHRG1, we cloned PyHRG1 between the 35S promoter and GFP in the plant expression vector 326-GFP (Fig. 4A). The recombinant PyHRG1-GFP construct was then introduced into tobacco (N. benthamiana) protoplasts and onion (A. cepa) epidermis cells. Fluorescence of the PyHRG1-GFP fusion protein was not detected in tobacco protoplasts (Fig. 4B). However, green fluorescence was predominantly observed in the cell wall region of the onion epidermis (Fig. 4C). These results demonstrate that PyHRG1 encodes a secretory protein located in the cell wall or extracellular matrix. In plants, secreted proteins play major roles in cell wall assembly and modification, as well as in responses to biotic and abiotic stresses (Wang et al. 2004). The plant cell wall determines cell size and shape through the mechanical control of cell expansion. Common responses to heat stress observed in the cell wall architecture include thickening and reduction in plasticity (Le Gall et al. 2015). Cell wall proteins, which mediate cell enlargement and expansion, include xyloglucan endo-β-transglucosylases/hydrolases, endo-1,4-β-D-glucanase, and expansins (Eklöf and Brumer 2010). Pectin-modifying enzymes, such as pectin methylesterase, also play a major role in controlling cell wall plasticity (Sénéchal et al. 2014). Previous studies have shown that some cell wall proteins are required for abiotic stress responses in plants (Choi et al. 2011, Wu et al. 2018). During secondary cell wall formation, monolignols (precursors of lignin) as well as cellulose and hemicelluloses are secreted into the cell wall space (Vanholme et al. 2010, Le Gall et al. 2015). Expression of such cell wall proteins can be altered in response to heat stress (Xu and Hwang 2018). Other cell wall-related genes also play a role in the acquisition of thermotolerance (Yang et al. 2006, Ha et al. 2007, Choi et al. 2013, Rienth et al. 2013). Yang et al. (2006) reported that the cell wall-related genes might play a role in the acquisition of thermotolerance in Chinese cabbage (Brassica rapa L.). Ha et al. (2007) also found that the cell wall protein, GSAP3, from Panax ginseng plays a role in abiotic stress tolerance. However, cell wall composition and consequent interactions with the environment and abiotic stressors can vary between different plant groups (Popper et al. 2014, Chen et al. 2020). Overall, further research is needed to gain an improved understanding of the role of the cell wall in heat tolerance at the physiological, genetic, and biochemical levels in red algae, including P. yezoensis.
CONCLUSION
We generated transcriptomes sequences from both the wild-type SG104 P. yezoensis and heat-tolerant mutant Gy500, and selected DEGs that were up- or downregulated in response to the heat stress condition and in the heat-tolerant mutant Gy500. In this study, we characterized a high-temperature response gene PyHRG1, which was downregulated under heat stress in SG104 and expressed at a low level in the heat-tolerant mutant Gy500. PyHRG1 encodes a novel red algae-specific secretory protein. Transformed C. reinhardtii cells overexpressing PyHRG1 showed retarded cell growth. These results indicate that PyHRG1 is involved in cell growth during heat stress. PyHRG1 is a novel gene found only in red algae and further studies are needed to understand the molecular function of PyHRG1. The transcriptome sequences obtained in this study, which include PyHRG1, will facilitate future studies to understand the molecular mechanisms involved in heat tolerance in red algae.