INTRODUCTION
GENERAL EYESPOT BIOLOGY
GALACTOLIPIDS IN EYESPOT-CONTAINING ALGAE
Chlorophyceae
D. acidophila galactolipids were observed to contain high relative amounts of 18:3/16:4 MGDG (38.4%), 18:3/16:3 MGDG (10.3%), 18:3/16:2 DGDG (27.3%), 18:3/16:1 DGDG (25.0%), and 18:3/16:0 DGDG (18.7%) (Della Greca et al. 1989). Thus, the fatty acids of these galactolipids resemble those expected for this algal class.
P. incisa, an alga within the class Trebouxiophyceae but still within the same phylum as Chlorophyceae, was described as containing relatively high amounts of 16:2(n-6; 20.8%), 16:3(n-3; 11.0%), 18:2(n-6; 31.4%), and 18:3(n-3; 18.5%) fatty acids as part of MGDG, and 16:0 (34.0%), 18:2(n-6; 31.0%) fatty acids as part of DGDG, all with unknown regiochemical distributions (Bigogno et al. 2002b). In addition, Bigogno et al. (2002a) observed P. incisa to produce relatively large amounts of 16:2(n-6; 9.6%), 16:3(n-3; 23.4%), 18:2(n-6; 15.3%), and 18:3(n-3; 32.6%) fatty acids as part of MGDG and 16:0 (19.2%), 18:2(n-6; 22.2%), 18:3(n-3; 26.1%), and eicosatetraenoic acid [20:4(n-6; 12.3%)] fatty acids as part of DGDG, all with unknown regiochemical distributions. While this alga is of the class Trebouxiophyceae, the major fatty acids and fatty acid biomarkers for this class are the same as those of Chlorophyceae (Taipale et al. 2013). These fatty acids generally match those expected of other green algae.
D. salina was reported as containing almost the entirety of its MGDG in the form of 18:3/16:4 (92.0%), and it also contained relatively high amounts of 18:3/16:3 (double bond positions undetermined) DGDG (46.3%), 18:3/16:0 DGDG (26.2%), and 18:3/16:4 DGDG (10.4%) (Lynch et al. 1983). Similar results were observed by Cho and Thompson (1987) in the thylakoid of D. salina as they reported relatively high amounts of 18:3/16:4 MGDG (92.5%), 18:3/16:3(n-3) DGDG (30.0%), 18:3/16:2 and 18:2/16:3(n-3) DGDG (16.7%), and 18:3/16:0 DGDG (25.6%).
The galactolipids of D. bardawil (synonymous with D. salina) were not described by Fried et al. (1982) in terms of fatty acid regiochemical (i.e., sn-1 vs. sn-2) positioning; however, they reported that MGDG was dominated by 18:3 (52.7%), 18:2 (16.2%), and 16:4 (29.7%) fatty acids. DGDG was dominated by 18:3 (25.7%), 18:2 (10.1%), and 16:0 (54.5%) fatty acids (Fried et al. 1982). These fatty acids also resemble those expected for Chlorophyceae.
D. tertiolecta was observed to possess relatively high amounts of 18:3/16:4 MGDG (63.1%) as well as 18:3/16:0 DGDG (16.6%) and 18:3/18:3 DGDG (12.3%) (Leblond et al. 2013). These fatty acids are generally characteristic of Chlorophyceae fatty acids.
The green alga C. reinhardtii was described as containing relatively large amounts of 18:3/16:4 MGDG (70%), 18:3/16:3 MGDG (16%), 18:3/18:3 DGDG (11%), 18:3/16:0 DGDG (18%), 18:2/16:0 DGDG (25%), and 18:1/16:0 DGDG (13%) (Giroud et al. 1988). As such, the fatty acids comprising chloroplast galactolipids are similar to those expected for this algal class. In addition, C. reinhardtii was observed containing relatively large amounts of MGDG containing 18:1 (15.2%), 18:2 (23.2%), and 18:3 (39.7%) fatty acids along with DGDG containing 16:0 (24.3%), 18:1 (10.6%), 18:2 (12.1%), and 18:3 (24.1%) fatty acids (Janero and Barrnett 1981); while the distributions of the MGDG and DGDG fatty acids are not noted, the fatty acids are comparable to those expected of the class Chlorophyceae.
Cryptophyceae
Cr. ovata (2 strains) was observed to contain relatively large amounts of 16:0 (16.0–16.3%), 18:3 (24.3–25.5%), 18:4 (21.3–21.4%), and 20:5 (15.0–16.4%) fatty acids (Mitani et al. 2017). These fatty acids closely resemble those expected for algae of this class. The fatty acid composition of Cr. acuta contained relatively large amounts of 16:0 (14.4%), 18:1 (13.4%), 18:3 (13.9%), and 18:4 (24.5%) fatty acids (Mitani et al. 2017).
The fatty acid composition of Ch. placoidea also contained relatively large amounts of 16:0 (24.0%), 18:1 (20.1%), 18:3 (17.3%), and 18:4 (9.8%) fatty acids (Mitani et al. 2017). These fatty acids are also typical of Cryptophyceae algae. Ch. caudata was described as containing relatively large amounts of 12:0 (12.1%), 14:0 (15.2%), 16:0 (19.6%), 18:1 (10.3%), 18:4 (13.4%), and 20:5 (9.4%) fatty acids (Mitani et al. 2017). Fatty acids of this species contain fatty acids characteristic to this class in addition to fatty acids [dodecanoic acid (12:0), tetradecanoic acid (14:0), and 18:1] not typically found in relatively large amounts.
The fatty acid composition of Ch. mesostigmata contained relatively large amounts of 16:0 (16.1%), 18:3 (16.3%), 18:4 (20.0%), and 20:5 (13.6%) fatty acids (Mitani et al. 2017). The fatty acid composition of Ch. coerulea (three strains) also contained relatively large amounts of 16:0 (13.3–22.8%), 18:3 (11.9–14.3%), 18:4 (17.6–28.9%), and 20:5 (10.5–14.7%) fatty acids (Mitani et al. 2017). These total fatty acids are characteristic of those expected for Cryptophyceae algae.
H. rufescens was observed as containing relatively large amounts of 16:0 (21%), hexadecenoic acid 16:1(n-7; 10%), 18:4(n-3; 17%), and eicosenoic acid 20:1(n-9; 14%) fatty acids, and small amounts of 18:3(n-3; 7%) and 20:5(n-3; 8%) fatty acids (Chuecas and Riley 1969). While Ch. mesostigmata and Ch. coerulea contained total major fatty acids characteristic of Cryptophyceae, H. rufescens contained unusual fatty acids [16:1(n-7; 10%) and 20:1(n-9; 14%)] that are not considered major fatty acids for this class. However, since Ch. caudata, which lacks an eyespot, also contained unusual fatty acids (12:0, 12.1%; 14:0, 15.2%; 18:1, 10.3%), differences in total fatty acid composition and possibly galactolipid composition could be a result of factors other than the presence or absence of an eyespot.
Euglenophyceae
E. gracilis was observed to possess relatively high amounts of 18:3/16:4 MGDG (29.0%), 20:4/16:4 MGDG (14.2%), and 18:3/16:3 DGDG (10.2%) (Craig et al. 2015).
L. acus was observed to produce high relative amounts of 18:3/16:4 MGDG (31.6%), 18:3/16:2 MGDG (13.9%), and 20:4/16:4 MGDG (14.9%) (Craig et al. 2015).
T. pomquetensis was observed to possess relatively high amounts of 16:0 (13.7%), 16:4(n-3; 14.8%), 18:4(n-3; 20%), and 20:5(n-3; 14.3%) fatty acids, in the absence of galactolipid data (McLachlan et al. 1999).
EYESPOT BIOLOGY IN DINOFLAGELLATES
GALACTOLIPIDS IN EYESPOT-CONTAINING DINOFLAGELLATES
Type A
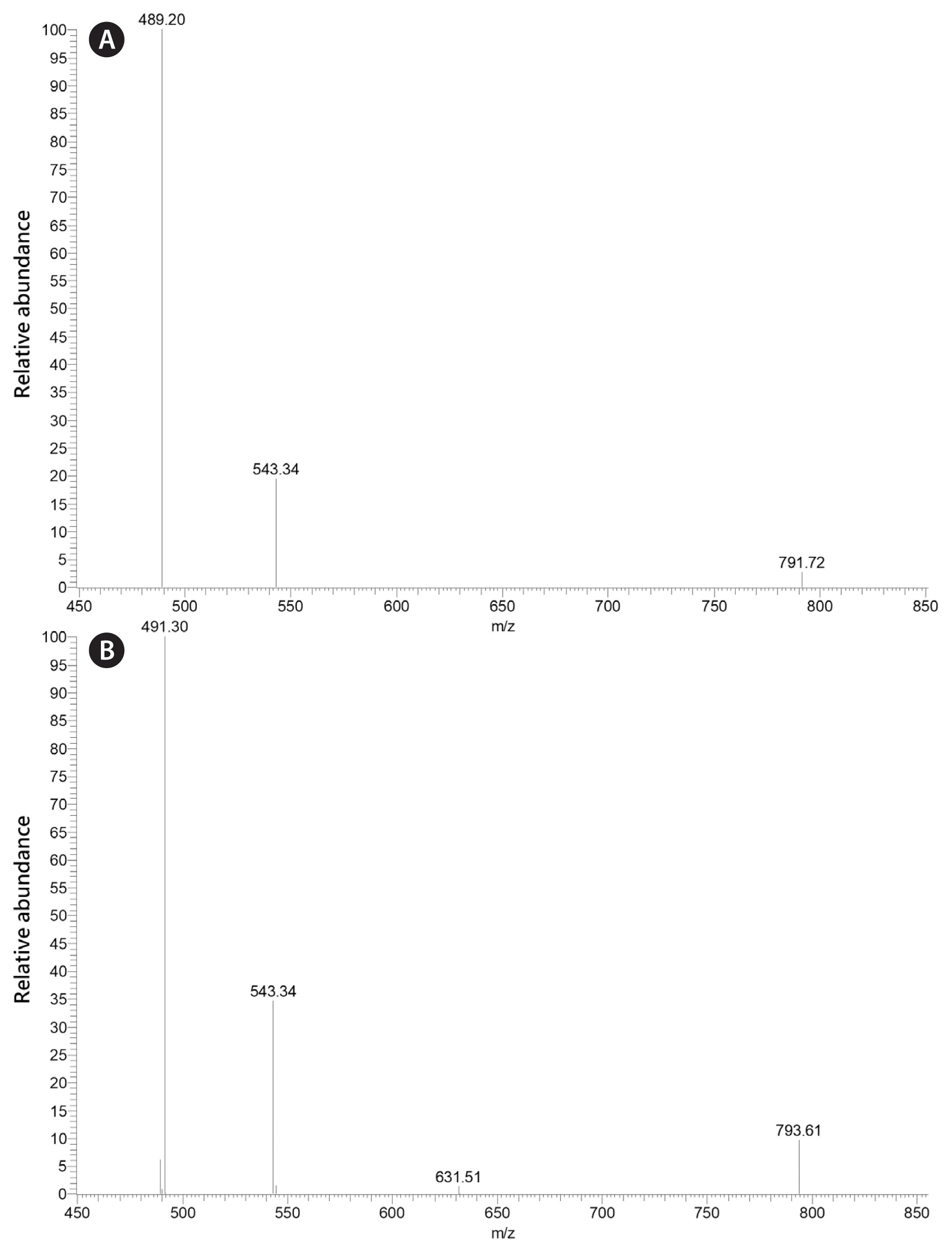
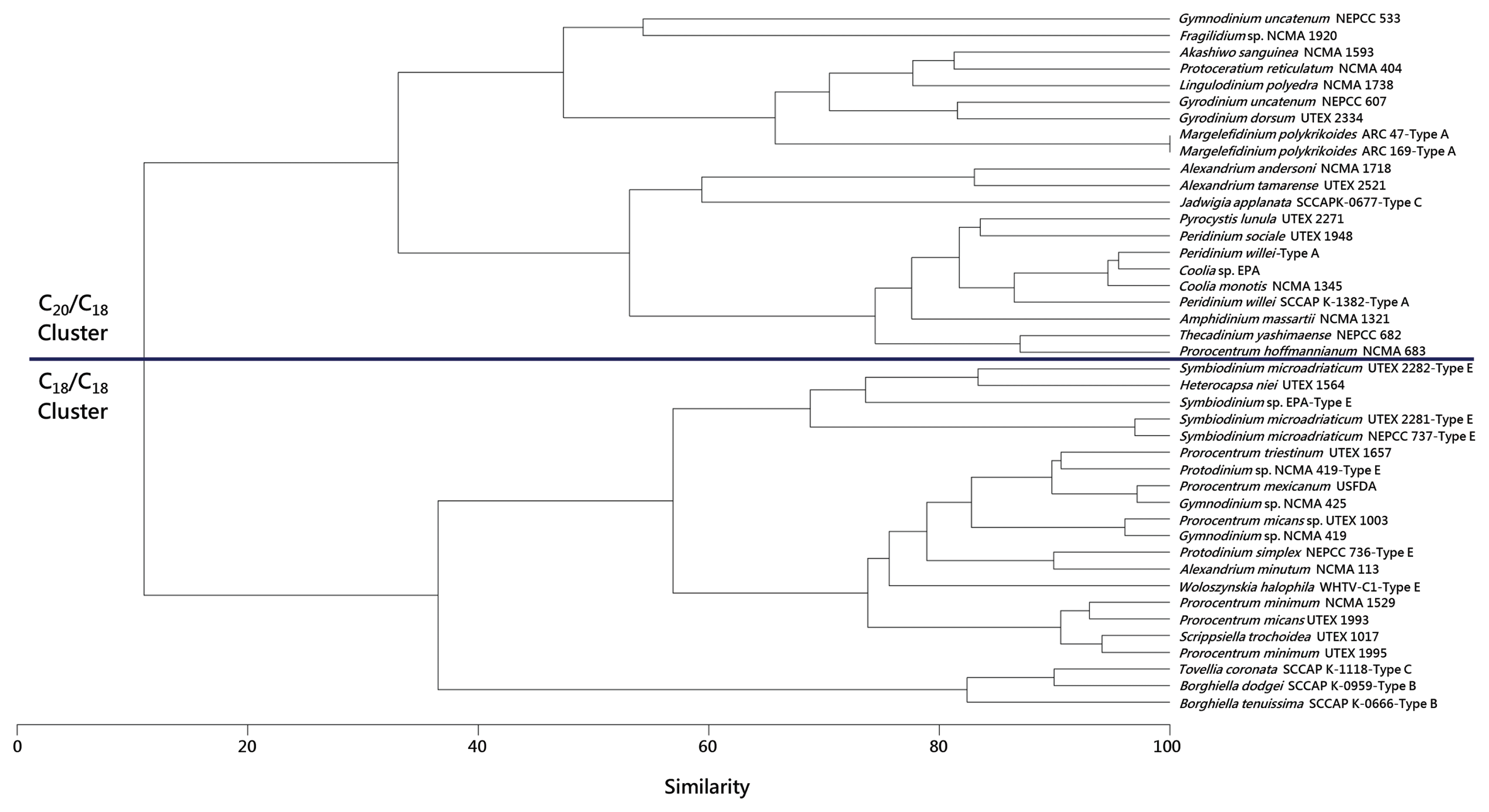
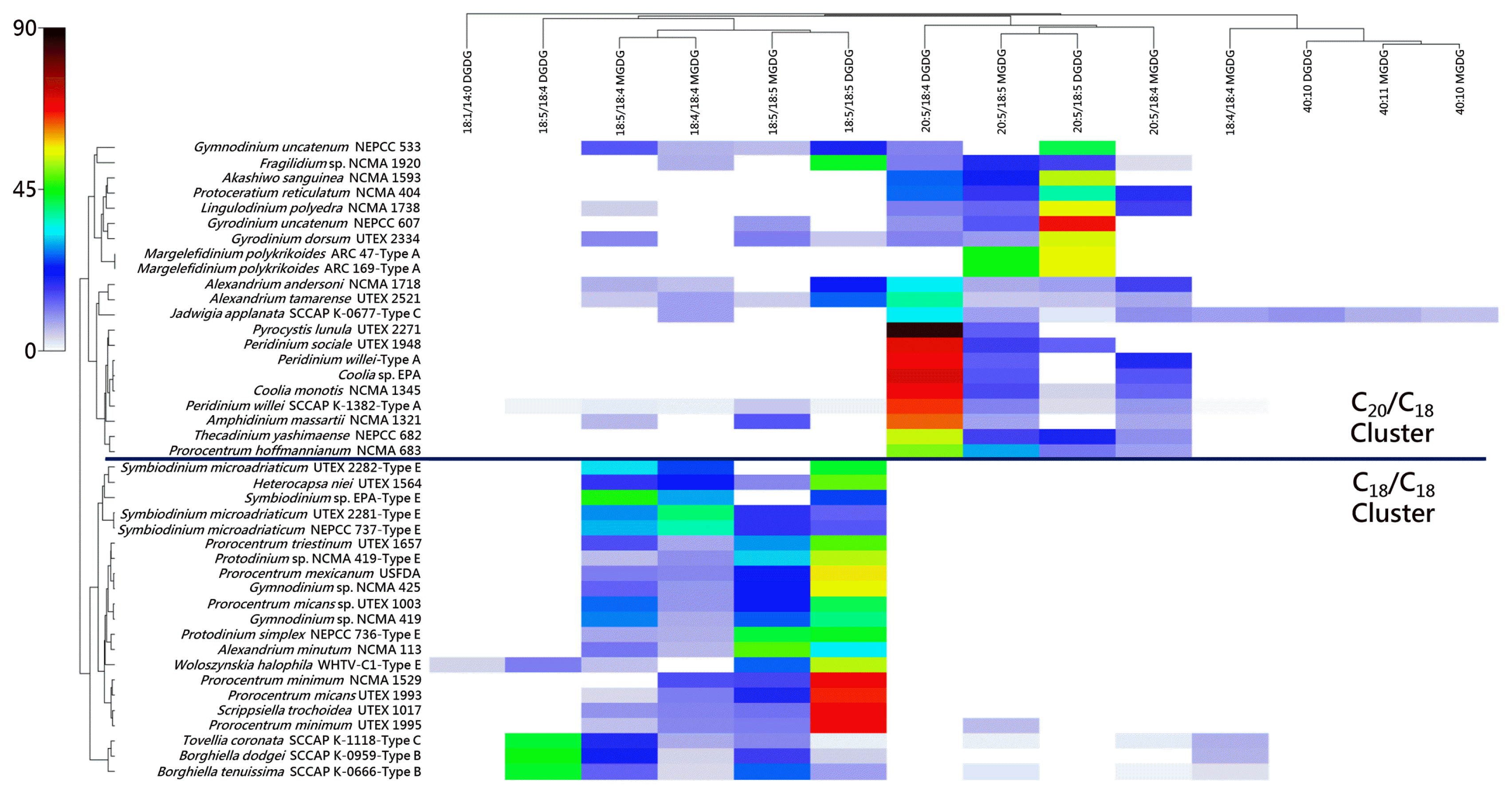
We have examined two isolates of M. polykrikoides (ARC 47 and ARC 169 from the Algal Resources Collection, Wilmington, NC, USA) according to the growth conditions and galactolipid characterization methodologies put forth in our previous papers on MGDG and DGDG in dinoflagellates, and these isolates were observed to possess 20:5/18:5 MGDG (m/z 817, approximately 45%) and 20:5/18:5 DGDG (m/z 979, approximately 55%). Both strains also possessed trace amounts of 18:5/18:5 MGDG (m/z 789) and 18:2/16:0 DGDG (m/z 939) yet showed no discernable difference in galactolipid composition from other dinoflagellates in the C20/C18 galactolipid cluster described earlier.
Per. willei was found by Gray et al. (2009b) to reside in the C20/C18 cluster with 20:5/18:5 MGDG, 20:5/18:4 MGDG, and 20:5/18:4 DGDG making up its galactolipid complement. The galactolipid composition of Per. willei as determined by Anesi et al. (2016) was marked by 38:10 MGDG, 38:9 MGDG, and 38:9 DGDG. Assuming that these are the 20:5/18:5 MGDG, 20:5/18:4 MGDG, and 20:5/18:4 DGDG, respectively, listed in their supplementary data, to match the results of Gray et al. (2009b), this would also place this particular isolate of Per. willei firmly within the C20/C18 cluster.
Type B
Type C
J. applanata was observed by Anesi et al. (2016) to possess high relative amounts of 38:10 MGDG, 38:9 MGDG, and 38:9 DGDG. These represent 20:5/18:5 MGDG, 20:5/18:4 MGDG, and 20:5/18:4 DGDG, respectively, thus placing J. applanata firmly in the C20/C18 cluster.
T. coronata was observed by Anesi et al. (2016) to possess high relative amounts of 36:9 MGDG and 36:9 DGDG, representing 18:5/18:4 MGDG and 18:5/18:4 DGDG, respectively. This places T. coronata firmly within the C18/C18 cluster.
Type D
K. foliaceum has been found to have forms of MGDG and DGDG with 20:5(n-3) in the sn-1 position and the multiply-unsaturated C16 fatty acids 16:3 and 16:2 (double bond positions undetermined) in the sn-2 position, in a manner identical to the pennate diatom, Navicula perminuta Grunow (Leblond and Lasiter 2009).
We have examined one isolate of D. baltica (ARC 210), and have observed it to possess 20:5/16:4 MGDG (m/z 791; relative percent abundance approximately 25%), 20:5/16:3 MGDG (m/z 793; relative percent abundance approximately 65%), and 20:5/16:2 DGDG (m/z 957; relative percent abundance approximately 10%).
Type E
P. simplex, Protodinium sp., and three isolates S. microadriaticum, along with one isolate of Symbiodinium sp., were found by Gray et al. (2009b) to reside in the C18/C18 cluster. All isolates had 18:5/18:5 MGDG, 18:5/18:4 MGDG, 18:5/18:4 DGDG, and 18:4/18:4 DGDG as their galactolipid complement.
A single isolate of W. halophila was examined by Gray et al. (2009a) and found to possess 18:5/18:5 MGDG, 18:5/18:4 MGDG, 18:5/18:5 DGDG, and 18:5/18:4 DGDG, along with a small relative percentage of 18:1/14:0 DGDG. Note that this isolate is cold-adapted and was grown at a lower temperature than the isolates examined in the Gray et al. (2009b) study, and that the increased level of unsaturation in 18:5/18:5 DGDG compared to the other type E eyespot dinoflagellates (which were not observed to possess this particular galactolipid) is likely a temperature-induced modulation.
The dinoflagellates listed are divided into the same C18/C18 and C20/C18 clusters originally observed by Gray et al. (2009b).
All type A eyespot taxa were located within the C20/C18 cluster, and all type B and type E taxa were located in the C18/C18 cluster.
Type C J. applanata was located in the C20/C18 cluster, while type C T. coronata was located in the C18/C18 cluster.