Physiological response of red macroalgae Pyropia yezoensis (Bangiales, Rhodophyta) to light quality: a short-term adaptation
Article information
Abstract
Light quality is a common environmental factor which influences the metabolism of biochemical substances in algae and leads to the response of algal growth and development. Pyropia yezoensis is a kind of economic macroalgae that naturally grows in the intertidal zone where the light environment changes dramatically. In the present study, P. yezoensis thalli were treated under white light (control) and monochromatic lights with primary colors (blue, green, and red) for 14 days to explore their physiological response to light quality. During the first 3 days of treatment, P. yezoensis grew faster under blue light than other light qualities. In the next 11 days, it showed better adaptation to green light, with higher growth rate and photosynthetic capacity (reflected by a higher rETRmax = 61.58 and Ek = 237.78). A higher non-photochemical quenching was observed in the treatment of red light than others for 14 days. Furthermore, the response of P. yezoensis to light quality also results in the difference of photosynthetic pigment contents. The monochromatic light could reduce the synthesis of all pigments, but the reduction degree was different, which may relate to the spectral absorption characteristics of pigments. It was speculated that P. yezoensis adapted to a specific or changing light environments by regulating the synthesis of pigments to achieve the best use of light energy in photosynthesis and premium growth and metabolism.
INTRODUCTION
The growth and metabolism of red macroalgae, a kind of photosynthetic autotrophs, were largely adjusted by the light environment in which they live. Mainly inhabiting coastal area (intertidal and subtidal area), red macroalgae confronts highly variable light qualities due to the periodical changes in water depth and turbidity momentarily, daily, and seasonally (Kirk 1994, Litjens et al. 1999, Morel et al. 2007). During the evolution, red macroalgae adapted to the heterogeneity with their special photosynthetic equipment and light absorption components (Talarico and Maranzana 2000, Hintz 2021).
Photosynthesis begins with the absorption of light by pigments molecules located in the thylakoid membrane (Johnson 2016). Due to the difference of pigments composition and content, high plants and alga of various phyla have their own absorption spectra, which are mainly centered on blue light (400–450 nm) and red light (650–700 nm), and furthermore, red alga have their unique absorption peak at green light (500–550 nm) (Barsanti et al. 2007). Chlorophyll a (Chl a) is an important pigment for light absorption in plants, and its maximum absorption peak is limited to 420–460 and 663 nm (Aronoff 1950). For red algae growing in drastically changing light environments, phycobiliprotein (PBP) is an important component of the light-harvesting complex, mainly including phycoerythrin (PE), phycocyanin (PC), allophycocyanin (APC), and linker proteins (Kursar and Alberte 1983). The binding of different PBPs ensures the ability of the red algae to collect energy over a wide range of wavelengths (490–650 nm) (Watanabe and Ikeuchi 2013) and allows efficient utilization of light energy (Hintz 2021), playing an important role in determining their ecological niche (Stomp et al. 2004) and adapting to variable light environment.
Under different light quality, the red macroalgae would change their synthesis of photosynthetic pigments. Previous studies showed that blue and green lights could induce the synthesis of PE and PC of Porphyra leucosticta (i.e., Pyropia leucosticta; Korbee et al. 2005), Meristotheca papulosa (Borlongan et al. 2020), Gracilaria birdiae (Barufi et al. 2015), and Halymenia floresii (Godínez-Ortega et al. 2008). The contents of PBP and Chl a decreased under red light in Pyropia haitanensis (Wu 2016). However, the Chl a synthesis was promoted by combining a certain proportion of red and blue lights in Gracilaria tikvahiae (Kim et al. 2015).
Photosynthesis and growth of red macroalgae are also affected by light quality. It was reported that under green light, red algae Merisrotheca papulosa (Borlongan et al. 2020) and Gracilaria lemaneiformis (i.e., Gracilariopsis lemaneiformis; Gong et al. 2020) had a higher maximum net photosynthetic rate. Wu (2016) reported that blue, green and fluorescent tubes gave rise to a higher photochemical activity and photosynthetic efficiency of Pyropia haitanensis and thus, to some extent, promoted its growth. Furthermore, compared with red and white light, blue light could improve the photochemical efficiency of Porphyra leucosticta with a higher value of Fv/Fm (Aguilera et al. 2000, Korbee et al. 2005). An interesting finding was that in Porphyra leucosticta, cells that acclimated to blue light had a higher potential for photosynthesis showed by a higher O2 production (Aguilera et al. 2000). Several studies showed that many species of red algae cultured under green light had the same or even higher growth rate than that under white light and generally higher than that under red light (Godínez-Ortega et al. 2008, Barufi et al. 2015, Gong et al. 2020).
Pyropia yezoensis is an important species naturally growing in the intertidal zone and widely cultivated in coastal area, exposing to variable light conditions (Sutherland et al. 2011). In this study, the growth rate, photosynthetic activity, and pigments content of P. yezoensis were investigated after being treated under different light qualities. The results may help understand the red macroalgal physiological response to varied light conditions in the intertidal environments. Moreover, this study would provide reference not only for exploring the photomorphogenesis and adaptation mechanism of algae to the external environment, but also for the improvement of cultivation methods for P. yezoensis.
MATERIALS AND METHODS
Algae materials
The thalli of P. yezoensis were obtained from a homozygous diploid conchocelis strain PYL 201306-440 isolated by the Laboratory of Algal Genetics and Breeding, Ocean University of China. The thalli were cultured in sterile sea water with Provasoli’s enriched seawater medium (Provasoli 1968) under the suitable growth temperature of 10 ± 1°C, the light intensity of 60 μmol m−2 s−1, and a 12 h light : 12 h dark photoperiod. The culture medium was renewed every three days.
Experimental treatments
Healthy thalli of P. yezoensis with bright red, smooth surface, length more than 10 cm, and width more than 2 cm were selected from cultured materials. Four discs of 8 mm diameter were cut from the middle part of each thallus and placed in darkness for 24 h. All 160 discs were randomly divided into four groups with 40 discs in each group, and were cultured in the instrument of MC-1000 Algae Culture and Online Monitoring System (PSI Photon Systems Instruments, Drasov, Czech Republic), with each treatment providing a kind of primary color light (blue, peaking at 470 nm; green, peaking at 540 nm; red, peaking at 640 nm) with the same peak light intensity of 60 μmol m−2 s−1 and a 12 h light : 12 h dark photoperiod. The white LED light (peaking at 450 nm and 550–600 nm) was set as the control group. Aeration provided water motion, and the temperature was kept at 10 ± 1°C water bath by IKA RC2 Basic Cooling Circulation System (IKA Works Guangzhou, Guangzhou, China). The P. yezoensis discs were cultured under different light qualities for 14 days and their areas (mm2) and chlorophyll fluorescence were measured on days 0, 3, 6, 9, 12, and 14. After the measurement on the last day, all the P. yezoensis discs were collected respectively and then frozen in liquid nitrogen after removing all surface moisture for determination of pigment contents.
Growth rate
Four groups of P. yezoensis discs were taken out, respectively, spread out on Petri dishes and taken photos by a single lens reflex camera. The areas were calculated by ImageJ (software (National Institutes of Health, Bethesda, MD, USA). The average daily growth rate (DGR) was assessed according to the formula DGR (% d−1) = [(At/ A0)1/t − 1] × 100% adapted from Yong et al. (2013), where A0 refers to the initial area and At refers to the area after t days.
Photosynthetic activity
Chlorophyll fluorescence was detected using the FluorCam MF800 Chlorophyll Fluorescence Imaging System (PSI Photon Systems Instruments). The P. yezoensis discs were acclimated to the dark for 15 min before being detected. The quenching curves were detected for measuring the maximum quantum yield of photosystem II (PSII; Fv/Fm), the quantum yield of PSII (ΦPSII), and non-photochemical quenching (NPQ). Fv/Fm was calculated from (Fm − Fo)/Fm, with ΦPSII from (F′m − Ft)/F′m, and NPQ from (Fm/F′m) −1 (Van Kooten and Snel 1990, Maxwell and Johnson 2000, Baker 2008). The rapid light curves (RLCs) were measured to assess the photosynthetic activity, by exposing them to six incremental steps of actinic light ranging from 39.6 to 977.5 μmol m−2 s−1 (photosynthetically active radiation [PAR]). Parameters of the RLCs were calculated by the following formulae adapted from Platt et al. (1980) and Ye et al. (2012):
Photosynthetic pigment contents
Each group of P. yezoensis discs was divided into three samples, and each sample’s wet weight was precisely weighed. The samples were ground with liquid nitrogen. The extraction of PE, PC, APC, and Chl a and the calculation of their contents were made according to Cao et al. (2021), and PBP content was the sum of PE, PC, and APC contents.
Spectral reflectance
Sixteen samples of P. yezoensis thalli were randomly selected to obtain hyperspectral images by hyperspectral camera (Specim IQ, Specim Ltd., Oulu, Finland). The spectral measurements were based on the line scanner and comprised the wavelength range of 400 to 1,000 nm. After the conversion of raw spectral radiance images to spectral reflectance images, the average reflectance of the region of interest for each sample was extracted by using Environment for Visualizing Images (ENVI) 5.1 software (Exiles Visual Information Solutions, Boulder, CO, USA).
Statistical analysis
Statistical analysis was performed using IBM SPSS Statistics version 25.0 (IBM Corp., Armonk, NY, USA). The growth, photosynthetic parameters, and pigments content were analyzed by one-way analysis of variance (ANOVA) with α = 0.05, followed by the t-test to compare the differences among treatments.
RESULTS
Growth rate
Four treatments showed varying degrees of growth under different light qualities (Fig. 1). It showed that P. yezoensis kept growing under different light qualities, and the difference between treatments was more and more obvious with the extension of treatment time. To show the response of P. yezoensis at the early stage of treatment and the adaptation after some time, the growth rates of the first 3 days and the 3rd to 14th days were selected for comparison (Fig. 1B). During the first 3 days of treatment, the P. yezoensis under blue light had a significantly higher growth rate than other monochromatic light, as well higher than white light although not significantly (p > 0.05). The growth rates of the P. yezoensis grown under green and red lights were significantly lower than that under white light (p < 0.05). However, with the extension of treatment time, the growth rates decreased under white and blue light treatments. During the period from the 3rd day to the 14th day of treatment, the growth rate of the P. yezoensis grown under green light was significantly higher than that under white and blue light (p < 0.05), which of under red light was not significantly different with that under white and green light (p > 0.05), and which of under blue light was the lowest than other light qualities (p < 0.05).

The areas (A) and average daily growth rate (DGR % d−1) (B) of Pyropia yezoensis grown under different light qualities. Different lowercase letters indicate a significant difference in the same period (p < 0.05).
Photosynthetic activity
Fig. 2 illustrates the maximum quantum yield (Fv/Fm), the quantum yield of PSII (ΦPSII), and NPQ derived from the quenching curves of the P. yezoensis grown under different light qualities at different treatment time, and the results of the 3rd and 14th day were chosen for detailed comparison. All treatments’ Fv/Fm values of increased gradually with the extension of treatment time. The red light treatment had the highest value throughout the treatment time (Fig. 2A). The comparison in Fig. 2D showed that the Fv/Fm values of P. yezoensis grown under red and blue lights were significantly higher than those under white light on the 3rd day of treatment (p < 0.05). And there was no significant difference between the green light treatment and the white light treatment (p > 0.05). On the 14th day of treatment, all treatments showed an increase in Fv/Fm values. There was no significant difference in Fv/Fm values among P. yezoensis grown under blue, green, and white lights (p > 0.05). The Fv/Fm value of the red light treatment was higher than those of the other treatments (p < 0.05). The ΦPSII values of each treatment fluctuated at different processing times (Fig. 2B). As shown in Fig. 2E, on the 3rd day of treatment, the ΦPSII values of P. yezoensis grown under green and white lights were higher than those under blue and red lights (p < 0.05). But on the 14th day of treatment, the ΦPSII values of the white and green light treatments decreased, and the differences among all the treatments were insignificant (p > 0.05). The difference in NPQ among treatment groups was gradually noitceable with the extension of treatment time (Fig. 2C). The NPQ variation trends of white, blue, and red light treatments were stable. In contrast, the value of the green light treatment showed some fluctuation. As shown in Fig. 2F, on the 3rd day of treatment, the NPQ value of P. yezoensis grown under red light was higher than that under white light (p < 0.05), and the NPQ values of the remaining three treatments were not significantly different (p > 0.05). On the 14th day of treatment, the NPQ value of the red light treatment was still higher. But there was no significant difference in NPQ value between any two treatments (p > 0.05) except that the red light treatment was significantly higher than the green light treatment (p < 0.05).
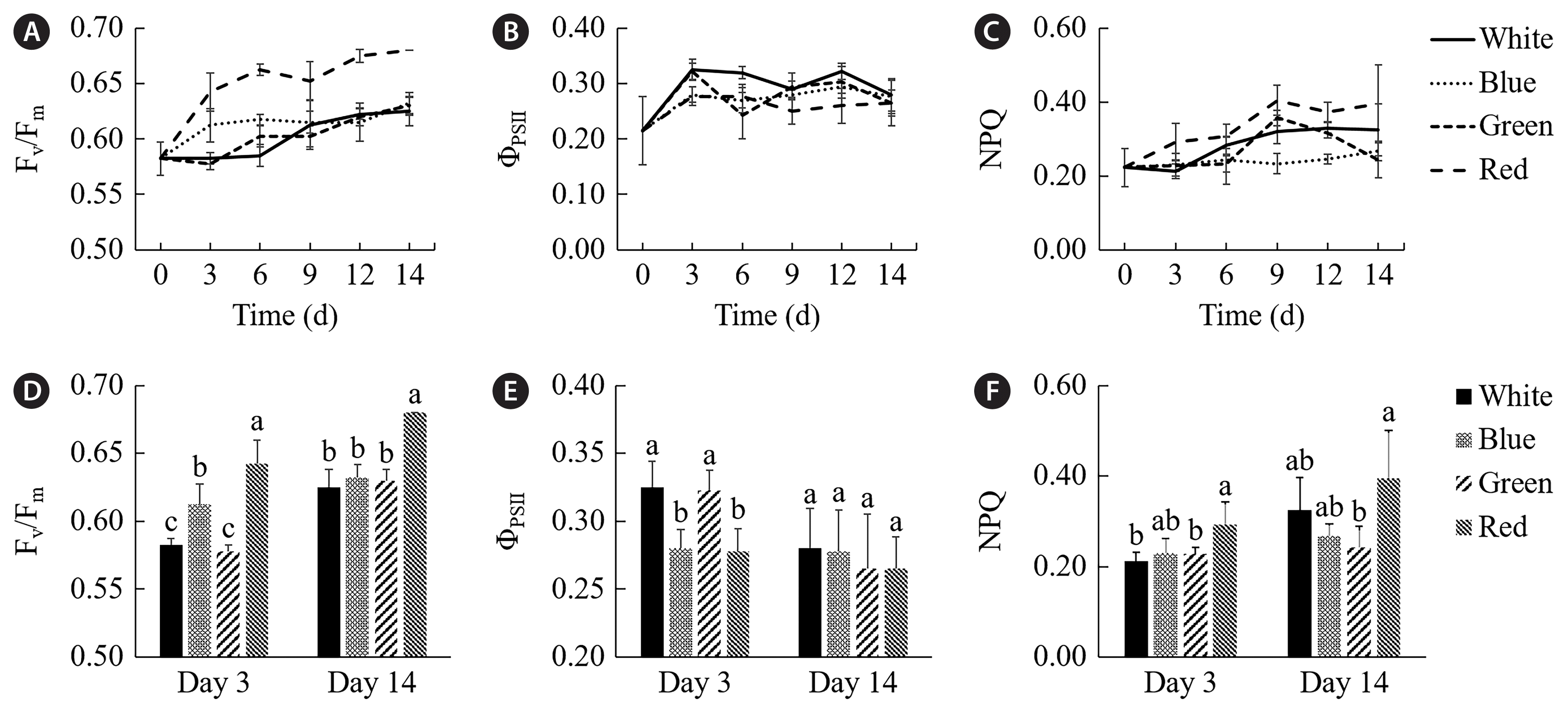
Quenching curve parameters (A & D, Fv/Fm; B & E, ΦPSII; C & F, non-photochemical quenching [NPQ]) of Pyropia yezoensis grown under different light qualities. Different lowercase letters indicate a significant difference in the same period (p < 0.05).
Fig. 3 illustrates the efficiency of electron transport (α), maximum relative electron transfer rate (rETRmax), and minimum saturation irradiance (Ek) derived from the RLCs of the P. yezoensis grown under different light qualities at additional treatment time, and the results of the 3rd and 14th day were chosen for detailed comparison. With the extension of the treatment time, the α value of each treatment had an upward trend, and the red light treatment showed the most significant increase range (Fig. 3A). In Fig. 3D, on the 3rd day of treatment, the α values were not significantly different among the four treatments (p > 0.05). On the 14th day of treatment, blue and red lights increased the α value, and the α value of the red-light treatment was higher than that of the white light treatment (p < 0.05). The rETRmax and Ek showed differences along with time under different light qualities (Fig. 3B & C). In Fig. 3E & F, rETRmax and Ek showed a similar pattern: on the 3rd day of treatment, there was little difference in rETRmax and Ek among the four treatments; on the 14th day of treatment, white and green lights increased the α value slightly.

Rapid light curves parameters (A & D, α; B & E, maximum relative electron transfer rate [rETRmax]; C & F, Ek) of Pyropia yezoensis grown at different light qualities on days 3 and 14. Different lowercase letters indicate a significant difference in the same period (p < 0.05).
Photosynthetic pigment contents
The pigment contents of P. yezoensis grown under different light qualities were summarized in Fig. 4. The content of each pigment was the highest at the white light. Compared with white light, blue light significantly reduced all the pigments contents (p < 0.05); green light was detrimental to the synthesis of PE and APC (p < 0.05) but had little effect on PC and Chl a (p > 0.05); and red light significantly limited the synthesis of PE, APC, and Chl a (p < 0.05), but did not affect substantially PC (p > 0.05).

Phycoerythrin (PE), phycocyanin (PC), allophycocyanin (APC), and chloroplyll a (Chl a) contents of Pyropia yezoensis grown under different light qualities. Different lowercase letters indicate a significant difference (p < 0.05).
According to Nakajima and Ueda (1999), the light-harvesting pigment to chlorophyll content ratio was essential for microalgal photosynthetic productivity. So, the PBP / Chl a ratio was selected to illustrate the effect of light quality on the pigment content ratio of P. yezoensis (Fig. 5). The PBP / Chl a ratio of the green light treatment was the lowest among all the treatments (p < 0.05); blue light treatment showed the highest PBP / Chl a ratio (p < 0.05); and there was no significant difference between the red and white light treatments (p > 0.05).
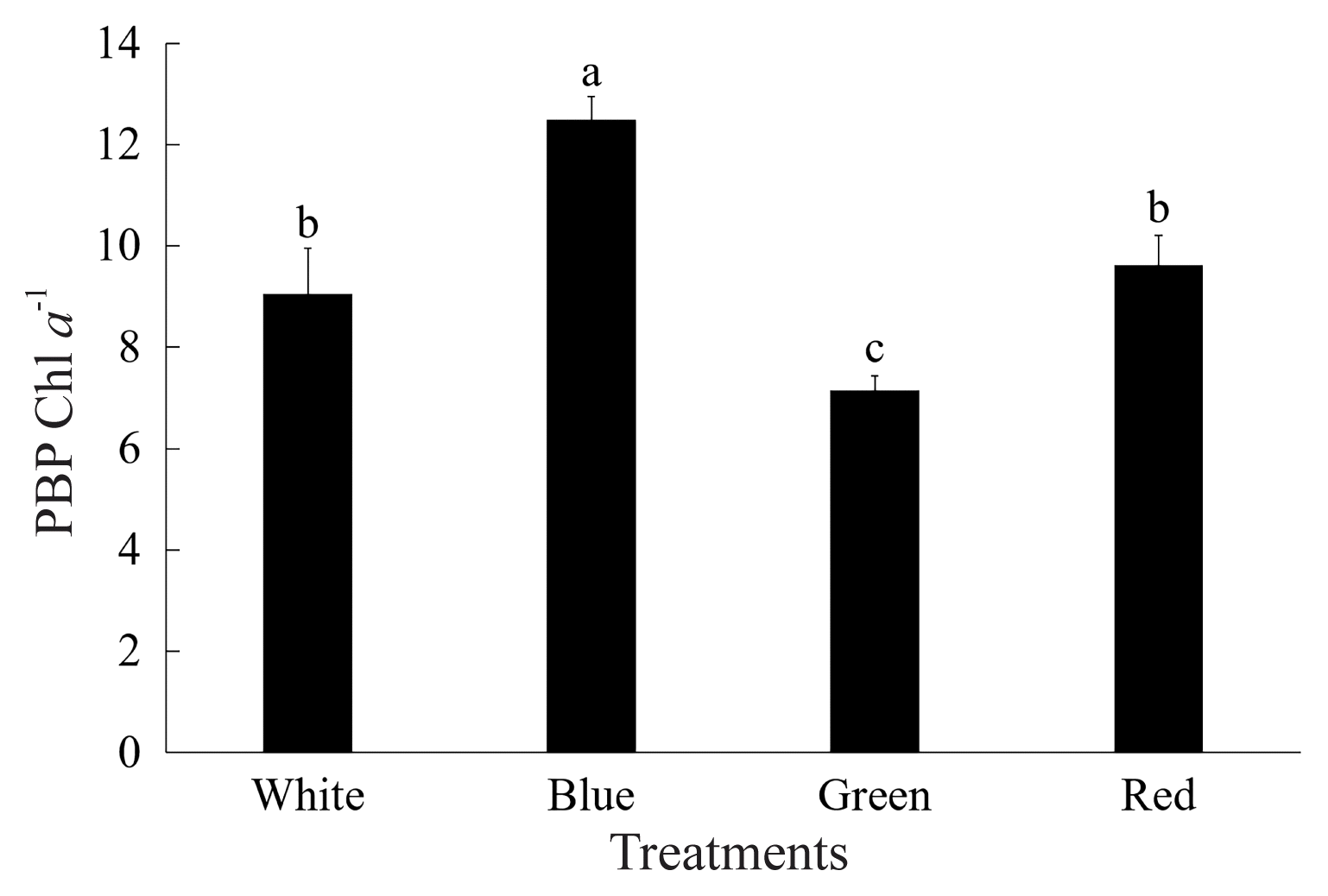
The phycobiliprotein (PBP) / chloroplyll a (Chl a) ratio of Pyropia yezoensis grown under different light qualities. Different lowercase letters indicate significant difference (p < 0.05).
Spectral reflectance
Fig. 6 illustrates the average spectral reflectance of P. yezoensis from 400 to 1,000 nm. The typical absorption wavelengths of P. yezoensis were distributed in the visible spectral region, around 440, 480, 570, 620, and 680 nm (marked by the dotted lines).

The average spectral reflectance of Pyropia yezoensis from 400 to 1,000 nm. The solid line is the mean and the range is given by grey bounding.
DISCUSSION
This study showed that different light qualities affected the growth of P. yezoensis, and in return P. yezoensis adapted to specific light attributes. Previous researches showed that the response of red macroalgal growth to light quality varied among species. Godínez-Ortega et al. (2008) reported that the growth rate of Halymenia floresii under green, blue, and red lights was higher than that under white light during the first 7 days of treatment, but from the 7th to 21st days, the growth rate of Halymenia floresii under blue light was inhibited compared with that under white light. Besides, it has been reported that the negative effect of blue light on the growth of Gracilaria tikvahiae was more pronounced than that of monochromatic green or red light (Kim et al. 2015). Conversely, research on Pyropia haitanensis showed that blue light was more efficient in promoting growth rate than red light (Wu 2016). In the present study, the growth rate of P. yezoensis under blue light was the highest during the first 3 days, even slightly higher than that grown under white light. But with the extension of the treatment time, the P. yezoensis gradually showed a growth advantage under green light and red light, which was similar to the results of Godínez-Ortega et al. (2008). Throughout the treatment period, the P. yezoensis could maintain a relatively stable growth rate under green and red lights, indicating that it had a better adaptation to green and red lights.
Different wavelengths and intensities of incident light could impact photosynthetic activity and photosystem adaption to light quality (Dietzel et al. 2008, Belkov et al. 2019). In this study, the photosynthetic activity of P. yezoensis showed a complex response to light quality. The Fv/Fm and ΦPSII indicated that during the first 3 days, P. yezoensis had a higher photosynthetic potential under blue and red lights. However, the practical PSII operating efficiency represented by ΦPSII was better when P. yezoensis thalli were under green and white lights. With the extension of treatment time, the degree of response of Fv/Fm in P. yezoensis under white, blue, and green lights was the same, but the Fv/Fm value of P. yezoensis under red light remained higher. Research has shown that a brief illumination of red light could cause an increased chlorophyll fluorescence yield, even if a very low intensity was used (Butler 1962). The changes of ΦPSII indicated that the practical PSII operating efficiency among the treatments tended to be consistent with the extension of treatment time. But this variation was erratic, possibly due to differences in the samples selected at each measurement.
NPQ is a parameter that indicates the ability of chloroplasts to dissipate excess excitation energy as heat when light absorbed by the photosynthetic apparatus surpasses the ability of alga for light utilization in photochemistry and protects alga themselves from photo-inhibition (Müller et al. 2001, Townsend et al. 2018, Bhagooli et al. 2021). In the present study, the higher NPQ value occurred in P. yezoensis under red light, indicating that even there was a higher maximum quantum yield (Fv/Fm) under red light, the light energy under this condition was not fully utilized, more entering the NPQ pathway and inducing photoprotection.
The RLC provides a reliable assessment of photosynthetic activity even where the light is rapidly fluctuating (Schreiber et al. 1997, White and Critchley 1999). In the present study, with the extension of treatment time, the parameters α, rETRmax and Ek under each treatment gradually manifested noticeable differences from their similarity at the beginning. P. yezoensis had better photosynthetic capacity (Ralph and Gademann 2005) under green and white lights indicated by the higher rETRmax values, in contrast to the lower electron transport efficiency shown by the lower α values, and thus resulted in higher minimum saturation irradiance (Ek). The photosynthetic pattern is similar to sun-adapted plants (Coe and Lin 2018). Besides, lower rETRmax and Ek values as well as higher α values were measured in red and blue lights, similar to the photosynthetic feature of shade-adapted plants (Reiskind et al. 1989, Coe and Lin 2018). Similarly, Gong et al. (2020) reported that the values of Pmax (reflecting the same meaning as rETRmax) were higher in green and white LEDs for red macroalgae Gracilaria lemaneiformis. The above photosynthetic characteristics indicated that P. yezoensis had better light adaptability and higher light use efficiency under green and white lights (Ralph and Gademann 2005, Coe and Lin 2018).
Photosynthesis begins with the light absorption by pigment molecules located in the thylakoid membrane (Johnson 2016). The energy of the photons absorbed by PE (λmax = 498 and 565 nm), PC (λmax = 620 nm), APC (λmax = 652 nm) (Grossman et al. 1995, Adir 2005), and Chl a (λmax = 431 and 663 nm) (Goedheer 1969, Duppeti et al. 2017) is funneled to the reaction center of the photosystems (Gantt 1981, Kursar and Alberte 1983). Algal photosynthetic pigments respond to the light environment in terms of content (López-Figueroa and Niell 1990, Wang et al. 2020, Latsos et al. 2021). From the results of this study, it can be found that light quality affects the synthesis of each pigment content to varying degrees. The PE content of P. yezoensis grown under blue light (peaking at 470 nm) was the highest among all monochromatic light treatments, while green light (peaking at 540 nm) and red light (peaking at 640 nm) had a less adverse effect on the synthesis of PC and APC. MacColl (1998) pointed out that the phycobilisomes of cyanobacteria formed complementary chromatic adaptation through changes in phycobilisomes rods, that is, light at the absorption peak of the pigment induced its production and changed the structure of the phycobilisomes. For Chl a, the lights near the maximum spectral absorption peaks were more unfavorable for its synthesis. From the spectral reflectance, it can be seen that the wavelengths of visible light absorption in P. yezoensis were mainly around the absorption peaks of PBPs and Chl a but did not precisely match their respective absorption peaks. The overlap of absorption spectra of different pigments may cause this difference.
However, the contents of all pigments under monochromatic light treatments were lower than that under white light treatments. The research of Lafarga-De la Cruz et al. (2006) stated that the reduction of pigment content is considered a process of autoregulation of the photosynthetic apparatus to reach a balance between the gain of light and the demand of energy necessary for growth. Besides, research suggested that a lower light-harvesting pigment content could result in higher photosynthetic productivity (Nakajima and Ueda 1999), which was consistent with the results of the present study. On the 14th day after treatment, the PBP / Chl a ratio of P. yezoensis grown under green light was the lowest. Correspondingly, the photosynthetic capacity (reflected by rETRmax) and growth rate of P. yezoensis under green light also showed a higher level. In another investigation, green light induced a higher photosynthetic activity and was harvested very efficiently by the PBPs (Borlongan et al. 2020). Therefore, the change of pigment content caused by specific wavelengths of light and the reasonable ratio among them have an essential effect on the efficient photosynthesis of algae, thus promoting growth.
In conclusion, P. yezoensis has specific physiological response under different light qualities. Among all the monochromatic light treatments, green light seems more favorable to the growth and photosynthesis of P. yezoensis. This adaptation to specific wavelengths of light enables red algae like P. yezoensis to have a growth advantage in regions that are mainly illuminated by blue-green light and is an important factor in determining their niche (Litjens et al. 1999, Morel et al. 2007, Hintz 2021). And such a response may be related to the fact that P. yezoensis regulates its pigments contents under this circumstance to succeed in a better adaptation. Therefore, for the mariculture of P. yezoensis, the light quality could be considered to obtain desirable agronomic features.
ACKNOWLEDGEMENTS
This study was financially supported by National Key R&D Program of China (2022YFD2400105, 2020YFD0900702, 2020YFA0607600). We thank Prof. Ik Kyo Chung (Pusan National University, Korea) and Christine Dupuy (La Rochelle University, France) for their improvement of this manuscript.
Notes
The authors declare that they have no potential conflicts of interest.