Ecophysiology of the kleptoplastidic dinoflagellate Shimiella gracilenta: II. Effects of temperature and global warming
Article information
Abstract
Water temperature affects plankton survival and growth. The dinoflagellate Shimiella gracilenta survives using the plastids of ingested prey, indicating kleptoplastidy. However, studies on the effects of water temperature on kleptoplastidic dinoflagellates are lacking. We explored the growth and ingestion rates of S. gracilenta as a function of water temperature. Furthermore, using data on its spatiotemporal distribution in Korean coastal waters during 2015–2018, we predicted its distribution under elevated temperature conditions of +2, +4, and +6°C. Growth rates of S. gracilenta with and without Teleaulax amphioxeia prey as well as ingestion rates were significantly affected by water temperature. Growth rates of S. gracilenta with and without prey were positive or zero at 5–25°C but were negative at ≥ 30°C. The maximum growth rate of S. gracilenta with T. amphioxeia was 0.85 d−1, achieved at 25°C, and 0.21 d−1 at 20°C without prey. The ingestion rate of S. gracilenta on T. amphioxeia at 25°C (0.05 ng C predator−1 d−1) was greater than that at 20°C (0.04 ng C predator−1 d−1). Thus, feeding may shift the optimal temperature for the maximum growth rate of S. gracilenta from 20 to 25°C. In spring and winter, the distributions of S. gracilenta under elevated temperature conditions were predicted not to differ from those during 2015–2018. However, S. gracilenta was predicted not to survive at some additional stations under elevated temperature conditions of +2, +4, and +6°C in summer or under elevated temperature conditions of +6°C in autumn. Therefore, global warming may affect the distribution of S. gracilenta.
INTRODUCTION
Water temperature frequently affects the survival, growth, and distribution of marine organisms (Loeng 1989, Gillooly 2000, Hiscock et al. 2004, Ok et al. 2018, Lee et al. 2019, Frölicher et al. 2020, Lim and Jeong 2021). Thus, there have been many studies on the effects of temperature on the ecophysiology of marine organisms (Beitinger and Fitzpatrick 1979, Thompson et al. 1992, Oliver and Palumbi 2011, Thomas et al. 2012, Lim et al. 2020, Kang et al. 2021). In addition, there have been various studies on predicting the distributions of several target marine organisms during climate change periods (Perry et al. 2005, Witt et al. 2010, Freeman et al. 2013, Jueterbock et al. 2013, Langer et al. 2013, Poloczanska et al. 2016). However, there are still many species whose responses to changes in water temperature should be explored.
Dinoflagellates are a major group of eukaryotic microorganisms in marine ecosystems that are found from the equator to the poles (Taylor et al. 2008, Jeong et al. 2021). They play diverse ecological roles, including primary producers, prey, predators, symbionts, and parasites (Coats 1999, Jeong 1999, Stat et al. 2008, Jeong et al. 2010, Montero et al. 2017, Ok et al. 2017, You et al. 2020a). Moreover, dinoflagellates frequently dominate natural assemblages, causing red tides or harmful algal blooms (Smayda and Reynolds 2003, Kudela and Gobler 2012, Jeong et al. 2013, 2015, 2021, Eom et al. 2021, Ok et al. 2021c). Therefore, changes in the distribution of dinoflagellate species can alter the structure and function of marine ecosystems. Dinoflagellates experience natural fluctuations in water temperature due to seasonal changes, and their distributions are known to be affected by changes in water temperature (Anderson and Rengefors 2006, Torres et al. 2019, Lee et al. 2020). Dinoflagellates grow the fastest at optimal water temperatures but die below or above a certain water temperature (e.g., Lim et al. 2019, Ok et al. 2019, Kang et al. 2020). The optimal water temperature and lower or upper temperature limits for the survival of dinoflagellates are species-dependent (e.g., Nielsen 1996, Laabir et al. 2011, Jeong et al. 2018, You et al. 2020b). Global warming elevates water temperature and may seriously affect the survival or distribution of dinoflagellates (Tunin-Ley et al. 2009, Kibler et al. 2015, Intergovernmental Panel on Climate Change 2021). Therefore, determining the optimal water temperature and lower and upper temperature limits for the survival of each dinoflagellate species is a critical step in understanding their ecophysiology. Furthermore, predicting the distribution of dinoflagellate species in the global warming period is important for predicting changes in the structure and function of marine ecosystems.
Shimiella gracilenta (previously Gymnodinium gracilentum) is a kleptoplastidic dinoflagellate that survives for approximately one month using the plastids of ingested prey cells (Skovgaard 1998, Ok et al. 2021b). This species can feed on prey species belonging to diverse taxa and is preyed on by some heterotrophic protists (Jakobsen et al. 2000, Ok et al. 2021a, Park et al. 2021). Furthermore, S. gracilenta was found at all 28 stations in Korea from 2015 to 2018 (Ok et al. 2021a). Thus, S. gracilenta may play diverse roles in marine ecosystems. In the late 21st century, Suh et al. (2016) predicted an increase in the surface air temperature by up to 5–8°C on the Korean Peninsula. Moreover, up to 4–5°C increase in seawater temperature surrounding the Korean Peninsula has been predicted (Kim et al. 2016). Thus, the distribution of S. gracilenta in Korean coastal waters may change in the future. To predict the future distribution and survival of S. gracilenta during the current global warming period, the growth rates of S. gracilenta under different temperature conditions and its lower and upper temperature limits for survival should be determined.
In the present study, we determined the growth rates of S. gracilenta SGJH1904 with and without the presence of the prey Teleaulax amphioxeia TAGS0202 and the ingestion rates as a function of water temperature. Furthermore, using data on its spatiotemporal distribution in Korean coastal waters during 2015–2018, we predicted its spatiotemporal distribution under elevated water temperatures that deviated +2, +4, and +6°C from the average for the period. The results of the present study provide a basis for understanding the effects of water temperature on the ecophysiological characteristics and distributions of S. gracilenta.
MATERIALS AND METHODS
Temperature effects on the growth and ingestion rates of Shimiella gracilenta
The growth and ingestion rates of S. gracilenta SGJH-1904 with and without added T. amphioxeia TAGS0202 at a single high prey abundance were measured at 5, 10, 15, 20, 25, 30, and 35°C. Clonal cultures of S. gracilenta SGJH1904 and T. amphioxeia TAGS0202 were established and incubated at 20°C as described in our previous study (Ok et al. 2021a). A culture of S. gracilenta growing on T. amphioxeia prey was transferred into a 10-L polycarbonate (PC) bottle. Four days later, the abundance of prey cells in the bottle was <10 cells mL−1. The dense culture in the bottle (ca. 70,000 cells mL−1) was transferred to seven 250-mL flasks. A dense culture of T. amphioxeia TAGS0202 (ca. 100,000 cells mL−1) was also transferred to each of the seven 250-mL flasks.
The target temperatures were established in seven temperature-controlled chambers. Prior to the experiment, the cultures were gradually acclimated to the target temperature for nine days to minimize the thermal shock on S. gracilenta and T. amphioxeia, following Lim et al. (2019), Ok et al. (2019), You et al. (2020b), and Kang et al. (2020) (Supplementary Fig. S1). During the preincubation period, 5-mL aliquots were collected from each flask at 3-day intervals and fixed with 5% Lugol’s solution to measure the abundance of S. gracilenta and T. amphioxeia. Finally, each culture of S. gracilenta or T. amphioxeia was placed in each chamber where the target temperature was established and incubated under a 14: 10 h light: dark cycle and 100 μmol photons m−2 s−1 of a light-emitting diode (LED; FS-075MU, 6500 K; Suram Inc., Suwon, Korea). However, in the preliminary test, all S. gracilenta and T. amphioxeia cells died at 35°C. Therefore, each culture of S. gracilenta and T. amphioxeia was acclimated at 25°C for 8 days and then at 30°C for 1 day, and these were used for the experiment at 35°C (Supplementary Fig. S1).
The initial single high prey abundance at which the growth and ingestion rates of S. gracilenta SGJH1904 on T. amphioxeia TAGS0202 were saturated was chosen (Ok et al. 2021a). Triplicate 38-mL flasks with predator-prey mixtures, predator-only controls (i.e., S. gracilenta only), and prey-only controls (i.e., T. amphioxeia only) were set up for each target temperature. The cell-free filtrates of S. gracilenta and T. amphioxeia were added to prey-only controls and predator-only controls, respectively, as described in our previous study (Ok et al. 2021a). Five milliliters of f/2 medium was added to all the flasks and filled with freshly filtered seawater. To determine the actual predator and prey abundances (cells mL−1) at the beginning of the experiment (Table 1), 5-mL aliquots were taken from each flask and fixed with Lugol’s solution. After subsampling at the beginning of the experiment, the flasks were refilled to capacity with freshly filtered seawater. The flasks were incubated for two days at each target temperature under a 14: 10 h light: dark cycle and 100 μmol photons m−2 s−1 of a LED. After a 2-day incubation, 10-mL aliquots were taken from each flask and fixed as described above.

Actual initial abundance (cells mL−1) of the predator Shimiella gracilenta SGJH1904 and prey Teleaulax amphioxeia TAGS0202
The specific growth rate (μ, d−1) of S. gracilenta SGJH-1904 was calculated using the following equation:
, where C0 and Ct represents the abundance of S. gracilenta at the beginning of incubation and after the 2-day incubation, respectively. The ingestion and clearance rates were calculated using the equation of Frost (1972) and the modified equation of Heinbokel (1978). The carbon content of T. amphioxeia was determined by Jeong et al. (2005).
The ingestion rates of S. gracilenta on T. amphioxeia at 30 and 35°C were not provided because the growth rates of S. gracilenta on T. amphioxeia at these water temperatures were negative, which could overestimate its ingestion rates.
Prediction of the distribution of Shimiella gracilenta under elevated water temperature
To explore the effects of warming on the survival of S. gracilenta, data on its spatiotemporal distribution and water temperature from 28 stations in Korean coastal waters from 2015 to 2018 were obtained from our previous study (Ok et al. 2021a). The water temperatures at each station in each season from 2015 to 2018 were averaged (+0°C). Then, 2, 4, and 6°C were added to the average water temperature at each station in each season (hereafter, +2, +4, and +6°C, respectively). To predict the presence or absence of S. gracilenta under elevated temperature conditions, the range of water temperatures for the survival of S. gracilenta, 1.7–26.4°C, was chosen from the results of our field observations (Ok et al. 2021a). Eight stations (Ansan, AS; Dangjin, DAJ; Mageompo, MGP; Taean, TA; Seocheon, SCN; Kunsan, KS; Buan, BA; and Mokpo, MP) were included in the West Sea of Korea, nine stations (Jangheung, JAH; Goheung, GH; Yeosu, YS; Kwangyang, KY; Tongyoung, TY; Masan, MS; Jinhae, JH; Dadaepo, DDP; and Busan, BS) in the South Sea of Korea, six stations (Ulsan, US; Pohang, PH; Uljin, UJ; Donghae, DH; Jumunjin, JMJ; and Sokcho, SC) in the East Sea of Korea, and five stations (Aewol, AW; Seogwipo, SGP; Wimi, WM; Seongsan, SS; and Gimnyeong, GN) in Jeju Island (Ok et al. 2021a).
Statistical analysis
To examine the effects of water temperature on the growth and ingestion rates of S. gracilenta SGJH1904, a one-way analysis of variance (ANOVA) with a post-hoc Tukey’s honestly significant difference (HSD) test was used. Prior to analysis, the normality and homogeneity of the data were tested using the Shapiro-Wilk test and Levene’s median test, respectively. To determine the differential effects of water temperature on the growth rates of S. gracilenta with and without prey, a multivariate analysis of variance (one-way MANOVA; Pillai’s trace test) was performed. Moreover, an independent sample t-test was conducted to test significant differences in the growth rates of S. gracilenta with and without prey. Statistical significance was set at p < 0.05. These analyses were performed using SPSS ver. 26.0 (IBM-SPSS Inc., Armonk, NY, USA).
RESULTS
Temperature effects on the growth rates of Shimiella gracilenta without prey
With increasing water temperatures, the growth rate of S. gracilenta SGJH1904 without added prey increased at 5–20°C but decreased at 25–35°C (Fig. 1). The maximum growth rate of S. gracilenta was achieved at 20°C. Furthermore, the growth rates of S. gracilenta without added prey at 5–25°C were zero or positive but negative at 30–35°C. The growth rates of S. gracilenta without prey ranged from −4.67 to 0.21 d−1. The growth rates of S. gracilenta without prey were significantly affected by water temperature (one-way ANOVA, F6, 14 = 3,089.2, p < 0.001) and were divided into four subsets (Tukey’s HSD post-hoc test, p < 0.05) (Fig. 1).
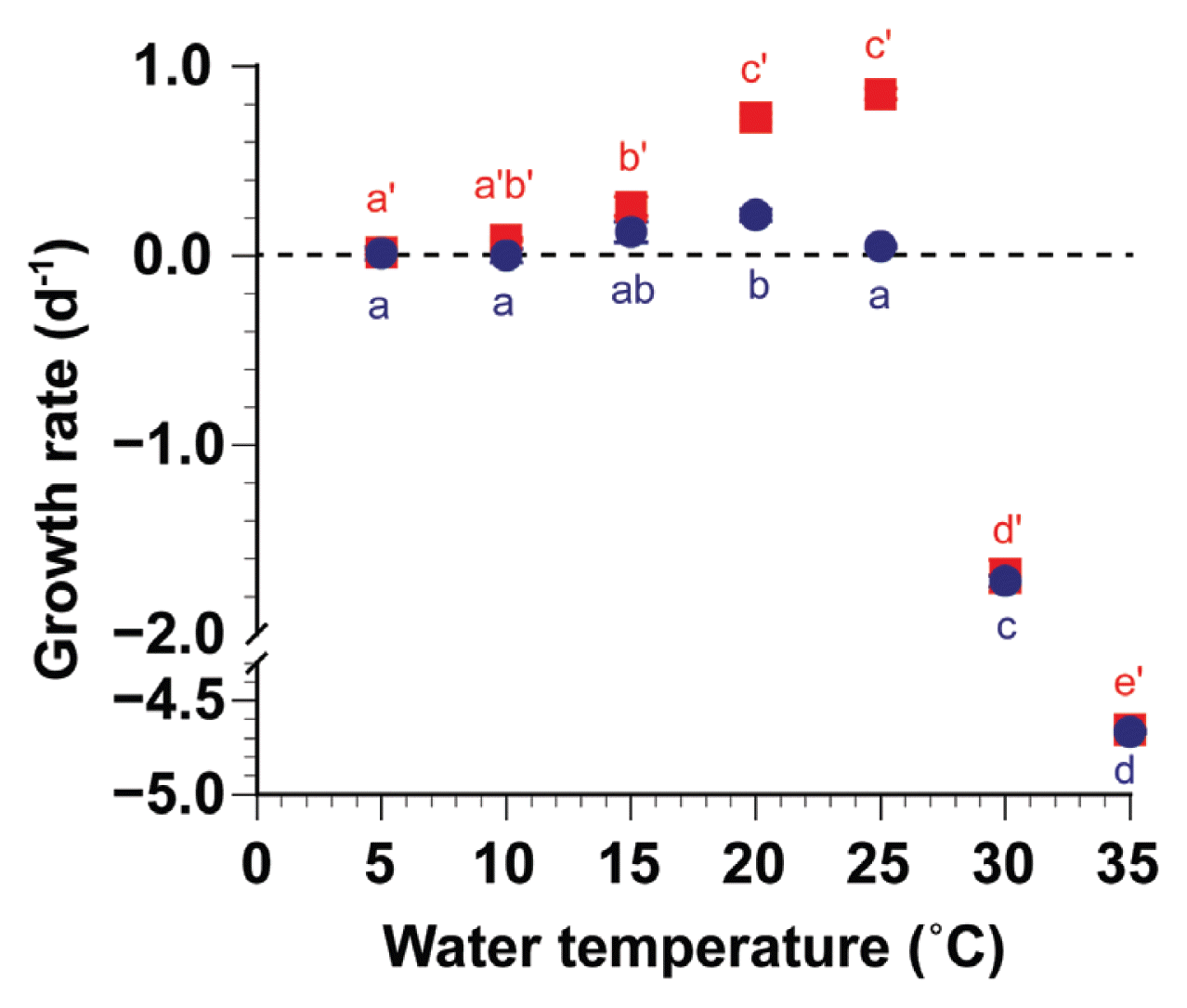
Specific growth rate of Shimiella gracilenta SGJH1904 with (red square) and without (blue circle) Teleaulax amphioxeia TAGS0202 as a function of water temperature. Significantly different subsets for the growth rate with (red letter with an apostrophe) and without prey (blue letter) were shown based on Tukey’s honestly significant difference post-hoc test after one-way ANOVA. Symbols represent treatment mean values ± standard error.
Temperature effects on the growth rates of Shimiella gracilenta with prey
With increasing water temperatures, the growth rate of S. gracilenta SGJH1904 with the added T. amphioxeia prey increased at 5–25°C but decreased at 30–35°C (Fig. 1). The maximum growth rate of S. gracilenta was achieved at 25°C. Furthermore, the growth rates of S. gracilenta with added T. amphioxeia prey were positive at 5–25°C but negative at 30–35°C. The growth rates of S. gracilenta with T. amphioxeia prey ranged from −4.66 to 0.85 d−1. The growth rates of S. gracilenta with T. amphioxeia prey were significantly affected by water temperature (one-way ANOVA, F6, 14 = 2,233.4, p < 0.001) and were divided into five subsets (Tukey’s HSD post-hoc test, p < 0.05) (Fig. 1).
Differential effects of water temperature between growth rates with and without prey
A MANOVA analysis revealed significant differences in the effects of water temperature on the growth rates of S. gracilenta SGJH1904 with and without prey (MANOVA, Pillai’s Trace = 1.93, F6, 14 = 60.6, p < 0.001) (Fig. 1). The growth rates of S. gracilenta with T. amphioxeia prey at 20°C and 25°C were significantly higher than those without added prey (one-tailed t-test, t4 = 13.3, p < 0.001 at 20°C; t4 = 27.9, p < 0.001 at 25°C).
Temperature effects on the ingestion rates of Shimiella gracilenta
With increasing water temperatures, the ingestion rate of S. gracilenta SGJH1904 on T. amphioxeia increased at 5–25°C (Fig. 2). The ingestion rate of S. gracilenta at 25°C, where its maximum growth rate with prey was observed, was 0.05 ng C predator−1 d−1 (2.9 cells predator−1 d−1). However, the ingestion rate of S. gracilenta at 20°C, where its maximum growth rate without prey was observed, was 0.04 ng C predator−1 d−1 (2.1 cells predator−1 d−1). The ingestion rates of S. gracilenta on T. amphioxeia were significantly affected by water temperature (one-way ANOVA, F4, 10 = 20.5, p < 0.001) and were divided into four subsets (Tukey’s HSD post-hoc test, p < 0.05) (Fig. 2).

Ingestion rate of Shimiella gracilenta SGJH1904 on Teleaulax amphioxeia TAGS0202 as a function of water temperature. Significantly different subsets for the ingestion rate were shown based on Tukey’s honestly significant difference post-hoc test after one-way ANOVA. Symbols represent treatment mean values ± standard error.
Prediction of the distribution of Shimiella gracilenta
In spring, the average water temperature during 2015–2018 at the 28 stations in Korea was 9.6–15.8°C, and S. gracilenta did not exist at 10 stations: one station in the West Sea, four stations in the South Sea, two stations in the East Sea, and three stations in Jeju Island (Fig. 3A). Under the +2, +4, and +6°C conditions in spring, the distribution of S. gracilenta was predicted not to differ from that during 2015–2018 spring seasons (Fig. 3B–D).
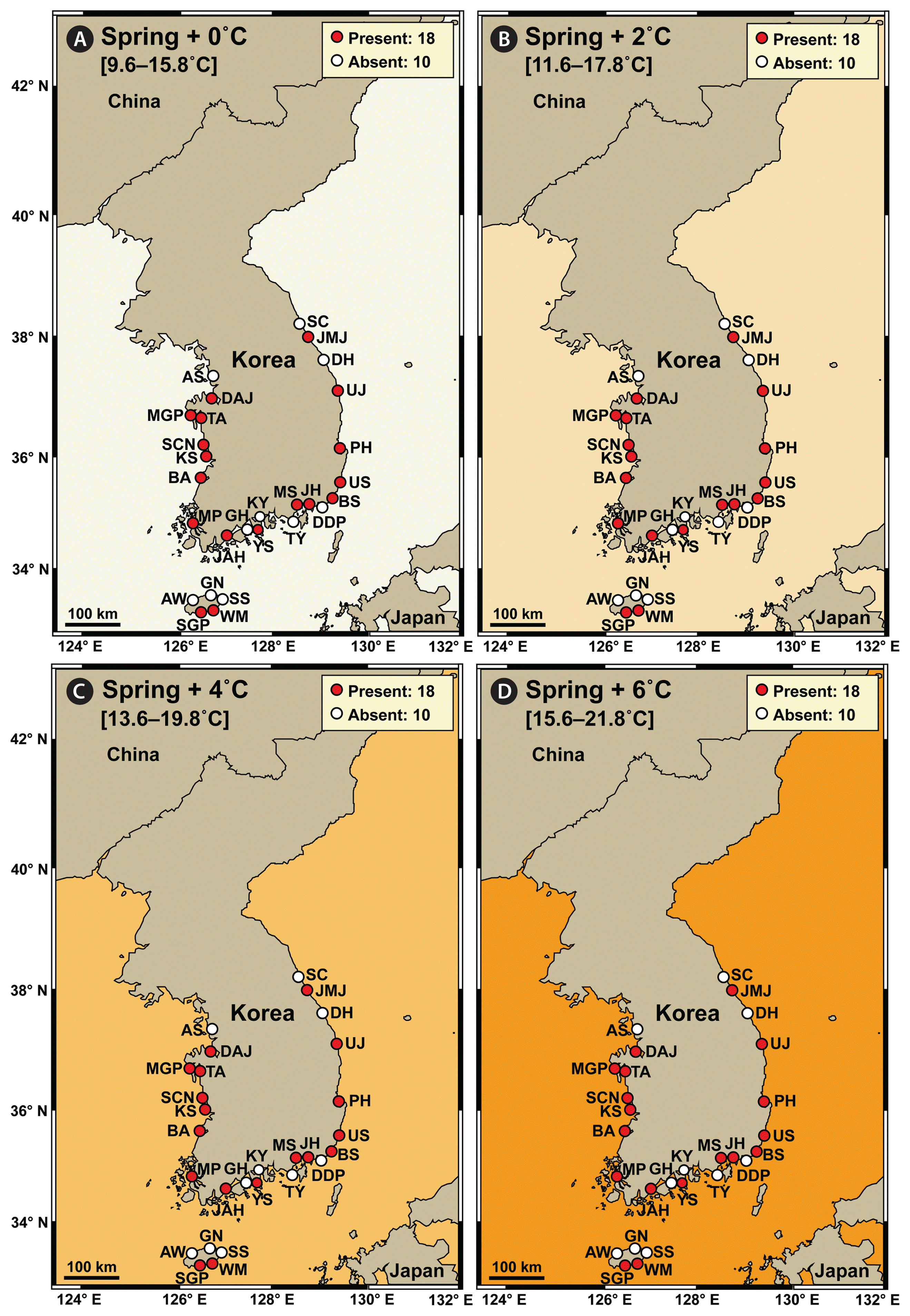
Map showing the results of a prediction of the nationwide distribution of Shimiella gracilenta in Korean coastal waters in spring under elevated water temperature conditions. The results were determined using the data on the distribution of S. gracilenta during 2015–2018 (Ok et al. 2021a). (A) Presence (red circles) or absence (white circles) of S. gracilenta at the average water temperature during the spring seasons of 2015–2018 (+0°C). (B–D) Prediction of presence or absence of S. gracilenta at the average water temperature plus 2°C (+2°C) (B), 4°C (+4°C) (C), and 6°C (+6°C) (D). Numbers in the box indicate the number of stations where S. gracilenta was (or was predicted to be) present or absent. The West Sea of Korea: Ansan (AS), Dangjin (DAJ), Mageompo (MGP), Taean (TA), Seocheon (SCN), Kunsan (KS), Buan (BA), and Mokpo (MP). The South Sea of Korea: Jangheung (JAH), Goheung (GH), Yeosu (YS), Kwangyang (KY), Tongyoung (TY), Masan (MS), Jinhae (JH), Dadaepo (DDP), and Busan (BS). The East Sea of Korea: Ulsan (US), Pohang (PH), Uljin (UJ), Donghae (DH), Jumunjin (JMJ), and Sokcho (SC). Jeju Island: Aewol (AW), Seogwipo (SGP), Wimi (WM), Seongsan (SS), and Gimnyeong (GN).
In summer, the average water temperature during 2015–2018 at the 28 stations in Korea was 19.1–25.7°C, and S. gracilenta did not exist at 10 stations: two stations in the West Sea, four stations in the South Sea, two stations in the East Sea, and two stations in Jeju Island (Fig. 4A). However, under the +2°C condition, S. gracilenta was predicted not to survive at 14 stations: five stations in the West Sea, five stations in the South Sea, two stations in the East Sea, and two stations in Jeju Island (Fig. 4B). Under the +4°C condition, S. gracilenta was predicted not to survive at 20 stations: eight stations in the West Sea, seven stations in the South Sea, two stations in the East Sea, and three stations in Jeju Island (Fig. 4C). Under the +6°C condition, S. gracilenta was predicted not to survive at 24 stations: eight stations in the West Sea, nine stations in the South Sea, two stations in the East Sea, and five stations in Jeju Island (Fig. 4D).
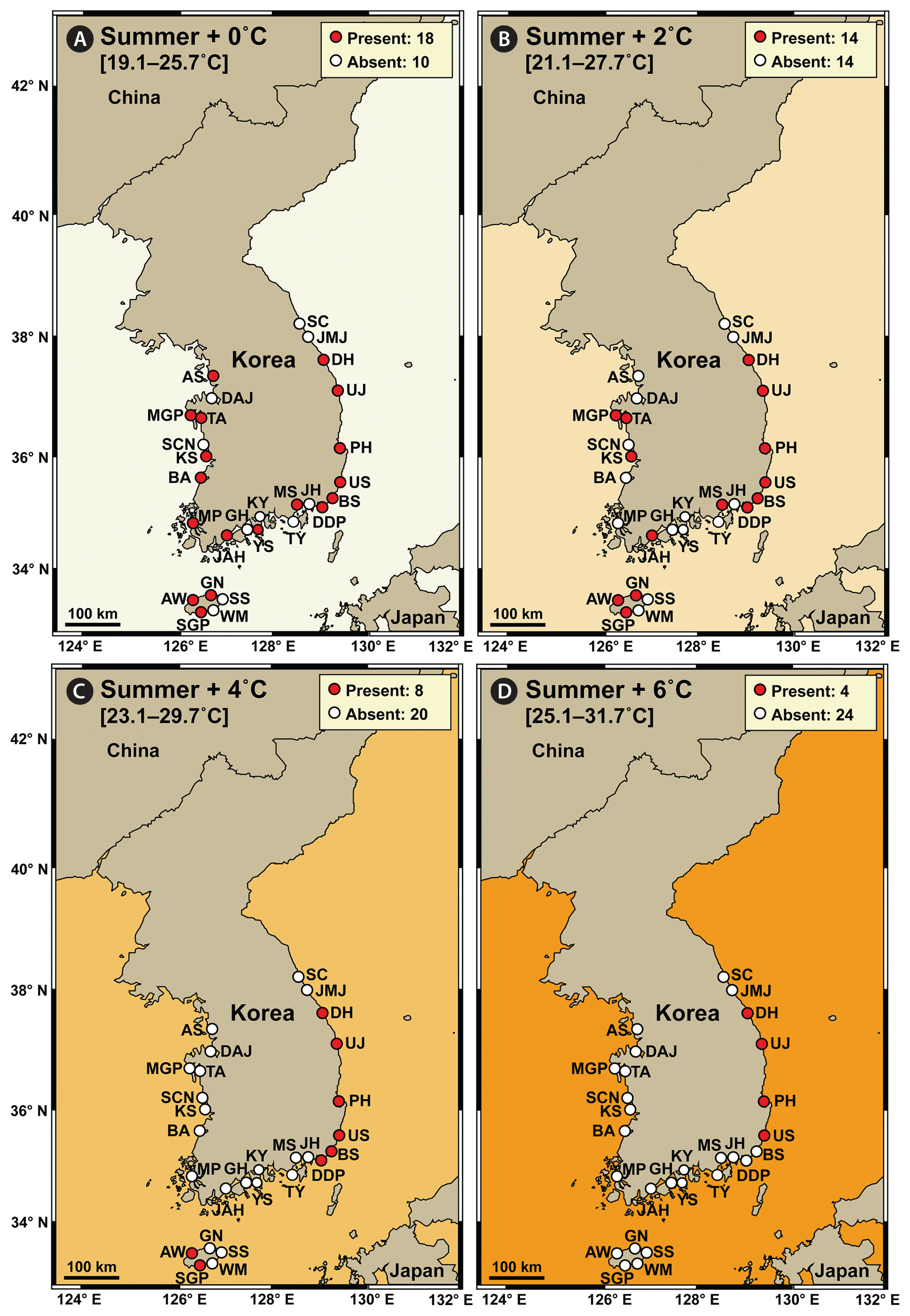
Map showing the results of a prediction of the nationwide distribution of Shimiella gracilenta in Korean coastal waters in summer under elevated water temperature conditions. The results were determined using the data on the distribution of S. gracilenta during 2015–2018 (Ok et al. 2021a). (A) Presence (red circles) or absence (white circles) of S. gracilenta at the average water temperature during 2015–2018 summer seasons (+0°C). (B–D) Prediction of presence or absence of S. gracilenta at the average water temperature plus 2°C (+2°C) (B), 4°C (+4°C) (C), and 6°C (+6°C) (D). Numbers in the box indicate the number of stations where S. gracilenta was (or was predicted to be) present or absent. The West Sea of Korea: Ansan (AS), Dangjin (DAJ), Mageompo (MGP), Taean (TA), Seocheon (SCN), Kunsan (KS), Buan (BA), and Mokpo (MP). The South Sea of Korea: Jangheung (JAH), Goheung (GH), Yeosu (YS), Kwangyang (KY), Tongyoung (TY), Masan (MS), Jinhae (JH), Dadaepo (DDP), and Busan (BS). The East Sea of Korea: Ulsan (US), Pohang (PH), Uljin (UJ), Donghae (DH), Jumunjin (JMJ), and Sokcho (SC). Jeju Island: Aewol (AW), Seogwipo (SGP), Wimi (WM), Seongsan (SS), and Gimnyeong (GN).
In autumn, the average water temperature during 2015–2018 at the 28 stations in Korea was 18.3–22.6°C, and S. gracilenta did not exist at 12 stations: three stations in the West Sea, five stations in the South Sea, two stations in the East Sea, and two stations in Jeju Island (Fig. 5A). Under the +2 and +4°C conditions in autumn, the distribution of S. gracilenta was predicted not to differ from that during 2015–2018 (Fig. 5B & C). However, under the +6°C condition, S. gracilenta was predicted not to survive at 19 stations: three stations in the West Sea, nine stations in the South Sea, two stations in the East Sea, and five stations in Jeju Island (Fig. 5D).
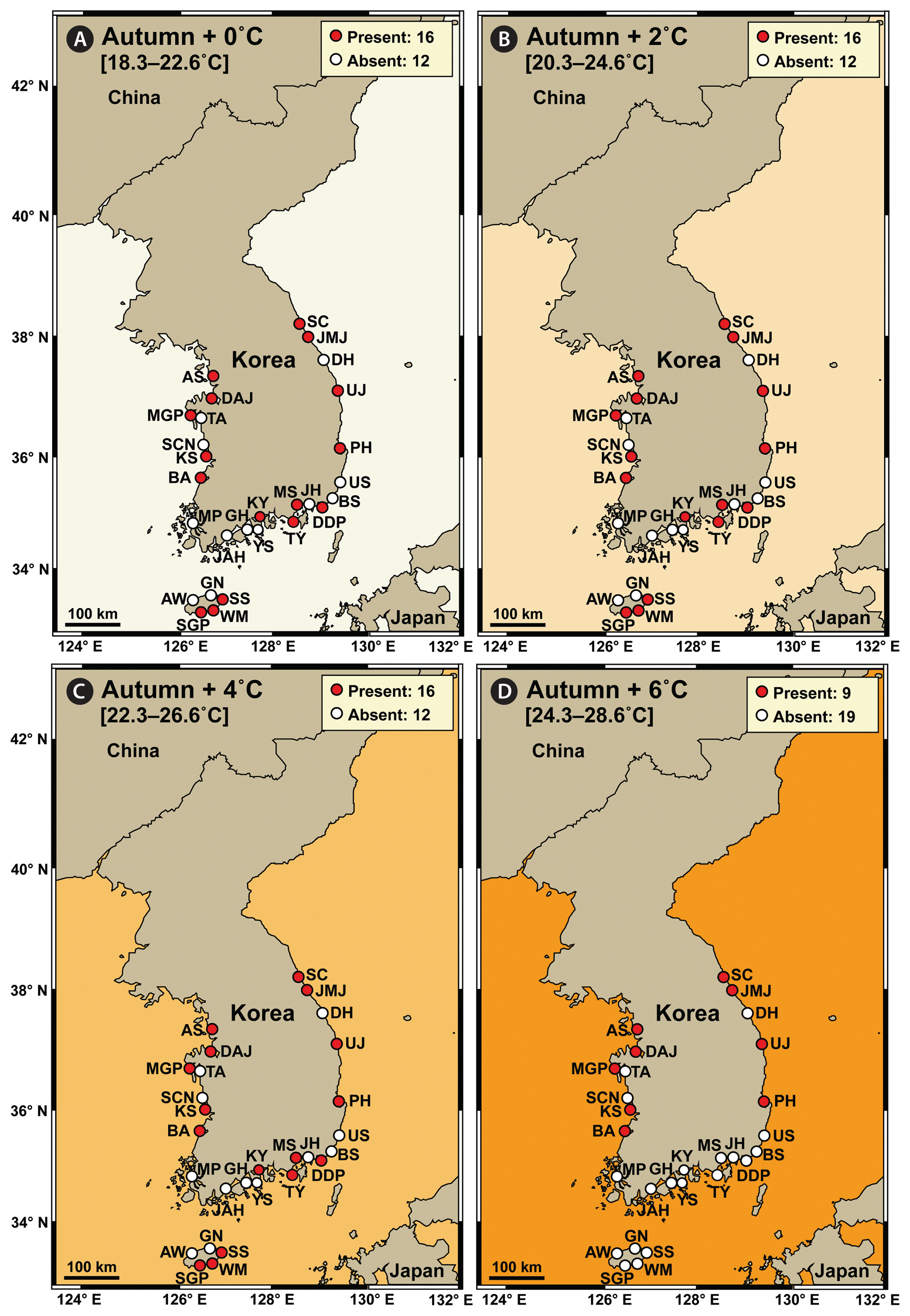
Map showing the results of a prediction of the nationwide distribution of Shimiella gracilenta in Korean coastal waters in autumn under elevated water temperature conditions. The results were determined using the data on the distribution of S. gracilenta during 2015–2018 (Ok et al. 2021a). (A) Presence (red circles) or absence (white circles) of S. gracilenta at the average water temperature during 2015–2018 autumn seasons (+0°C). (B–D) Prediction of presence or absence of S. gracilenta at the average water temperature plus 2°C (+2°C) (B), 4°C (+4°C) (C), and 6°C (+6°C) (D). Numbers in the box indicate the number of stations where S. gracilenta was (or was predicted to be) present or absent. The West Sea of Korea: Ansan (AS), Dangjin (DAJ), Mageompo (MGP), Taean (TA), Seocheon (SCN), Kunsan (KS), Buan (BA), and Mokpo (MP). The South Sea of Korea: Jangheung (JAH), Goheung (GH), Yeosu (YS), Kwangyang (KY), Tongyoung (TY), Masan (MS), Jinhae (JH), Dadaepo (DDP), and Busan (BS). The East Sea of Korea: Ulsan (US), Pohang (PH), Uljin (UJ), Donghae (DH), Jumunjin (JMJ), and Sokcho (SC). Jeju Island: Aewol (AW), Seogwipo (SGP), Wimi (WM), Seongsan (SS), and Gimnyeong (GN).
In winter, the average water temperature during 2015–2018 at the 28 stations in Korea was 4.2–16.2°C, and S. gracilenta did not exist at 15 stations: two stations in the West Sea, five stations in the South Sea, five stations in the East Sea, and three stations in Jeju Island (Fig. 6A). Under the +2, +4, and +6°C conditions in winter, the distribution of S. gracilenta was predicted not to differ from that during 2015–2018 winter seasons (Fig. 6B–D).
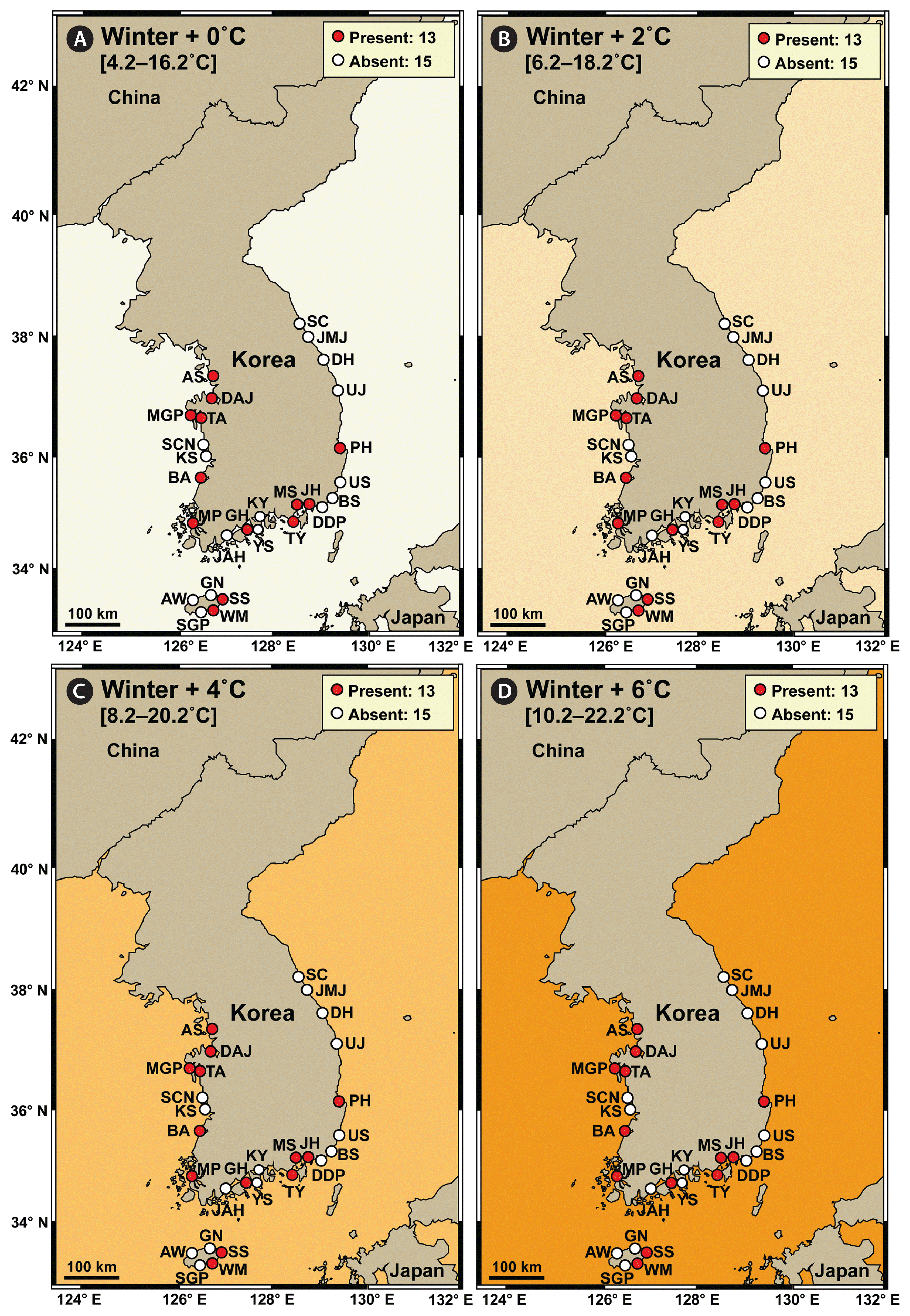
Map showing the results of a prediction of the nationwide distribution of Shimiella gracilenta in Korean coastal waters in winter under elevated water temperature conditions. The results were determined using the data on the distribution of S. gracilenta during 2015–2018 (Ok et al. 2021a). (A) Presence (red circles) or absence (white circles) of S. gracilenta at the average water temperature during 2015–2018 winter seasons (+0°C). (B–D) Prediction of presence or absence of S. gracilenta at the average water temperature plus 2°C (+2°C) (B), 4°C (+4°C) (C), and 6°C (+6°C) (D). Numbers in the box indicate the number of stations where S. gracilenta was (or was predicted to be) present or absent. The West Sea of Korea: Ansan (AS), Dangjin (DAJ), Mageompo (MGP), Taean (TA), Seocheon (SCN), Kunsan (KS), Buan (BA), and Mokpo (MP). The South Sea of Korea: Jangheung (JAH), Goheung (GH), Yeosu (YS), Kwangyang (KY), Tongyoung (TY), Masan (MS), Jinhae (JH), Dadaepo (DDP), and Busan (BS). The East Sea of Korea: Ulsan (US), Pohang (PH), Uljin (UJ), Donghae (DH), Jumunjin (JMJ), and Sokcho (SC). Jeju Island: Aewol (AW), Seogwipo (SGP), Wimi (WM), Seongsan (SS), and Gimnyeong (GN).
DISCUSSION
Using the culture of Shimiella gracilenta SGJH1904, the present study revealed that the growth rates were zero or positive at 5–25°C but negative at 30–35°C, regardless of the presence of added prey. Our previous study investigating the distribution of S. gracilenta at 28 stations in Korean coastal waters during the 2015 to 2018 period showed that the lowest water temperature at which S. gracilenta cells were detected was 1.7°C (Ok et al. 2021a). Thus, S. gracilenta may survive at water temperatures considerably lower than 5°C, which is the lowest water temperature tested in the present study. The highest water temperature at which S. gracilenta cells were found in Korean coastal waters during 2015–2018 was 26.4°C (Ok et al. 2021a). Therefore, the results from the field data were generally consistent with the experimental results of the present study.
At 20 and 25°C, the growth rates of S. gracilenta SGJH1904 with prey were 0.73 and 0.85 d−1, respectively, whereas those without prey were 0.21 and 0.05 d−1. Furthermore, the ingestion rates of S. gracilenta on prey at 20 and 25°C were 0.04 and 0.05 ng C predator−1 d−1, respectively. Thus, feeding elevated the growth rates of S. gracilenta to a greater extent at 25°C than at 20°C, and additionally, changed the optimal temperature for supporting the maximum growth rate.
The growth rates of T. amphioxeia, the prey species that supports the highest growth rate of S. gracilenta, were negative at ≥ 30°C (Kang et al. 2020). Therefore, water temperatures of ≥ 30°C may negatively affect the survival of S. gracilenta directly (i.e., physical damage) and indirectly (prey limitation).
The results of the present study showed that the distribution of S. gracilenta may be affected by global warming in summer and autumn months in Korean waters. According to our prediction, in summer, S. gracilenta is expected not to survive at some additional stations in the West and South Sea of Korea when the water temperatures are elevated by 2, 4, and 6°C above those temperatures recorded in 2015–2018, whereas S. gracilenta is expected to survive at most stations in the East Sea of Korea. In summer, warm currents such as the West Korea Coastal Current, the Tsushima Warm Current, and the Jeju Warm Current flow in the West Sea and South Sea of Korea and Jeju Island, while a southward cold current (the North Korea Cold Current) and a northward warm current (the Tsushima Warm Current) flows into the East Sea of Korea (Fig. 7) (Park et al. 2015, 2017). Therefore, the southward cold current can lower the water temperature in the East Sea of Korea in the summer and enable the survival of S. gracilenta here when it cannot survive in the West and South Seas of Korea and Jeju Island during this season.

Schematic maps of the surface currents in summer (A) and winter (B) in Korean waters, redrawn from Park et al. (2015, 2017).
During 1968–2004 in Korea, the water temperature at the surface increased by 0.975°C and at a depth of 10 m the temperature was elevated by 0.918°C (Jung 2008). This study demonstrated that warming affects the water temperature to a depth of 10 m. The average swimming speed of S. gracilenta has been reported to be 160 μm s−1 (Park et al. 2021). Theoretically, this dinoflagellate can migrate from the surface to a depth of 6 m for 10 h (Jeong et al. 2015). Therefore, the survival of S. gracilenta may be affected by warming sea water during the global warming period, despite its capability to descend to 6-m depths.
Shimiella gracilenta SGJH1904 was isolated from temperate coastal regions, while a sister species, the Ross Sea dinoflagellate W5-1 and RS-24, was isolated from Antarctica (Gast et al. 2006, Ok et al. 2021a, 2021b). The effects of water temperature on the survival of the Ross Sea dinoflagellate has not yet been reported. Thus, determining the effects of water temperature on the survival of the Ross Sea dinoflagellate and comparing this data with that of S. gracilenta SGJH1904 may provide insight into the adaptations and evolution of these two closely related species that live in contrasting habitats.
Numerous studies have been conducted on the effects of temperature and global warming on plankton communities, but fewer studies have examined the effects of temperature and global warming on populations of certain dinoflagellate species (Huertas et al. 2011, Yvon-Durocher et al. 2015, Jonkers et al. 2019, Lee et al. 2019, Benedetti et al. 2021). The abundance of dinoflagellate species can affect prey and predator populations (Smalley and Coats 2002, Kim et al. 2013, Yoo et al. 2013, Lee et al. 2017, Lim et al. 2017, Jang and Jeong 2020). Therefore, it is necessary that key species be assessed to increase the accuracy of models predicting the effects of temperature changes and global warming on the structure and function of marine ecosystems.
ACKNOWLEDGEMENTS
This research was supported by the National Research Foundation funded by the Ministry of Science and ICT (NRF-2020M3F6A1110582;NRF-2021M3I6A1091272; NRF-2021R1A2C1093379) award to HJJ.
Notes
The authors declare that they have no potential conflicts of interest.
SUPPLEMENTARY MATERIALS
Outline of the preincubation and experimental incubation periods of Shimiella gracilenta SGJH1904 (A) and Teleaulax amphioxeia TAGS0202 (B) (https://www.e-algae.org).