Platysiphon verticillatus and Arcticophycus glacialis: gen. et comb. nov. (Stschapoviales, Phaeophyceae) based on ecological considerations, life history, morphology, and ultrastructure
Article information
Abstract
Platysiphon verticillatus and Punctaria glacialis are synonymized by mainly on the basis of shared genes, nine identical sequences. Our hypothesis is that synonymy is erroneous. We posit the many differences of biological features between the two species, their ecological preferences, and occurrences do not support synonymy. Arcticophycus (nov. gen.) is described and contains the new combination Arcticophycus glacialis. Asexual, vegetative, life histories are proposed. Novel reproductive structures are described including previously unknown features of thallus development in the life history of both species. Discovery of the novel Platysiphon cyst, and dual sporangia types in Arcticophycus, is critical in establishing a probable strategy for survival through the Arctic winter for each annual species. Pl. verticillatus is erect, tubular, ovoid to circular in X sections, flaccid, non-parenchymatous, light brown, 10–15 cm high with distinctive photosynthetic hairs in clusters or whorled on the upper one third of the thallus. A. glacialis is erect, planar, robust, parenchymatous, dark brown and hairs absent. The Halosiphon clade is named, consisting of four genera: Halosiphon, Platysiphon, Arcticophycus, and Stschapovia. Occurrence of these genera is identified. Emended are the genus Platysiphon, the family Platysiphonaceae and the order Stschapoviales.
INTRODUCTION
To resolve the question of species identity we have attempted to bridge two compatible aspects of phycology, phylogeny and descriptive biological information. The former is demonstrable and useful in indicating broad taxonomic relationships (Jarvis et al. 2014), generic and higher, the latter often superior to recognize generic and specific concepts. Our data reinforces the integrity of two species, describes known portions of their life histories and posits their means of annual population over-wintering, previously undescribed for Platysiphon verticillatus, Arcticophycus glacialis and other Arctic annual species. Platysiphon verticillatus Wilce 1962 and Punctaria glacialis Rosenvinge 1910, arctic endemic thalloid brown algae, are assigned to synonymy by Kawai et al. (2015b). Support given for synonymy is mainly the co-occurrence (shared genes, with identical sequences) of nine DNA sequences, highly conservative chloroplast and mitochondrial genes (TAP, psaA, psaB, psbA, psbC, rbcL, cox1, and cox3), and nuclear ITS2 rDNA sequences (Kawai et al. 2015b, figs 1–3). Shared genes with identical sequences in algal, plant and animal taxa are commonplace (Campagna et al. 2017, Díaz-Tapia et al. 2017, Leonard 2017, Páll-Gergely 2017, Augyte et al. 2018), a central evolutionary theme of the biota in the fluctuating past environments of the N Atlantic Ocean and N Polar Sea.
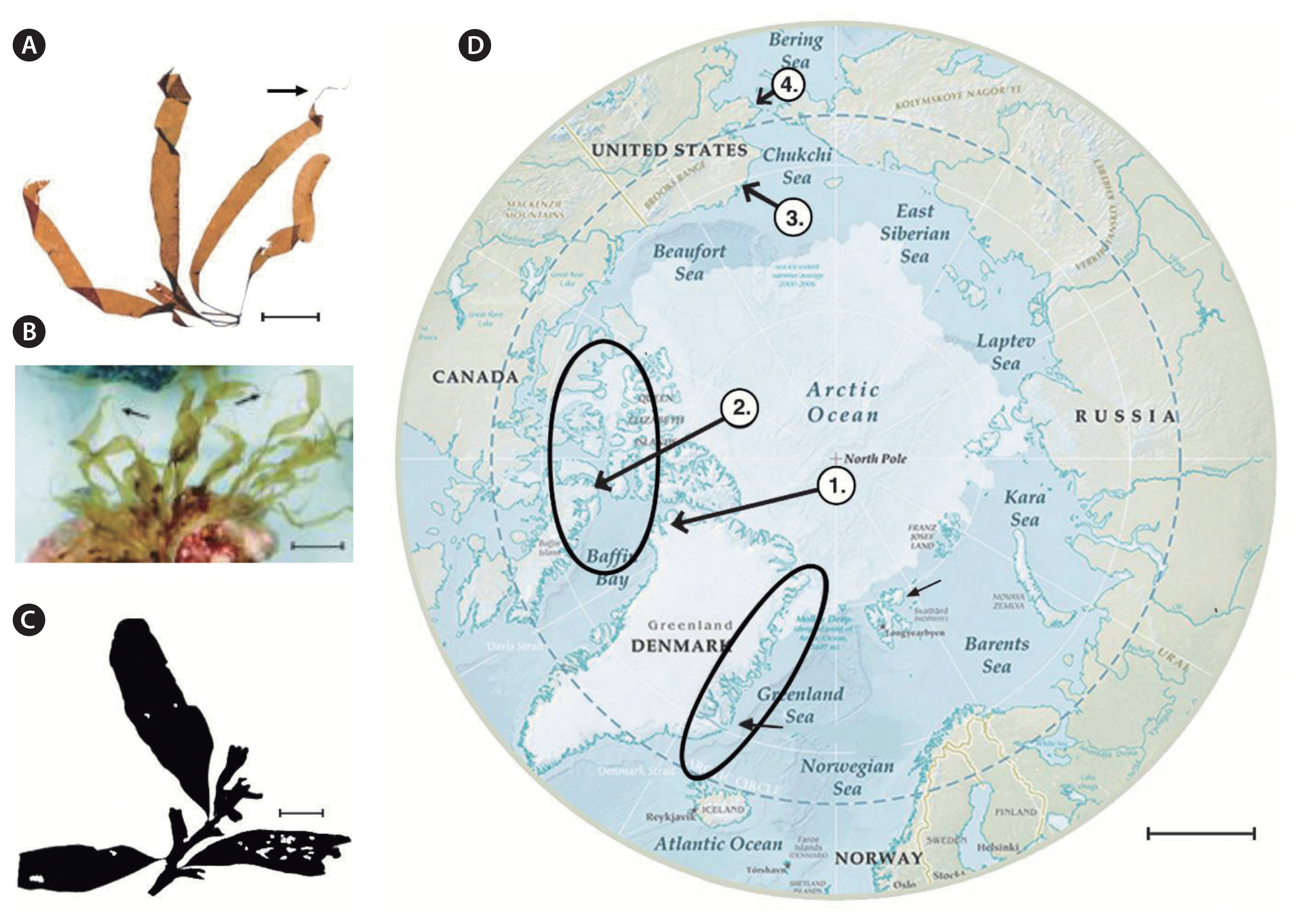
Platysiphon verticillatus and Arcticophycus glacialis: thalli size, contrast in morphology and a map of their known distributions. (A) Pl. verticillatus whole mount, vegetative, Sep 1983, ca. 3.0 m, Ragged Channel, Baffin Island, Nunavut, CAN., R.T.W., no. 7068-83, Univ. Mass. Herb.; see rat-tail (arrow). (B) Pl. verticillatus, attached thalli on rock/coralline substratum at 2–4 m, bay 10, Ragged Channel, Baffin Island, Nunavut, CAN., R.T.W., no. 7068-93, see rat tail (arrows) (preserved). (C) A. glacialis, whole mount, reproductive, epiphytic on Fucus distichus subsp. evanescens, Aug 1973, ca. 1.5 m, Melbourne Island, Nunavut, CAN., R.K.S. Lee, no. 9010, Nome, AK and CAN. Nat. Herb., Ottawa, CAN. (D) Polar projection map of Arctic Ocean and peripheral seas: Pl. verticillatus (arrows 1 & 2), Labrador, CAN, Saunders, collection site not shown, and A. glacialis (arrows 3 & 4), Nome, AK, Saunders, and within ellipses (modified U.S. Gov. Map). Scale bars represent: A, 3 cm; B & C, 10 cm; D, 350 km.
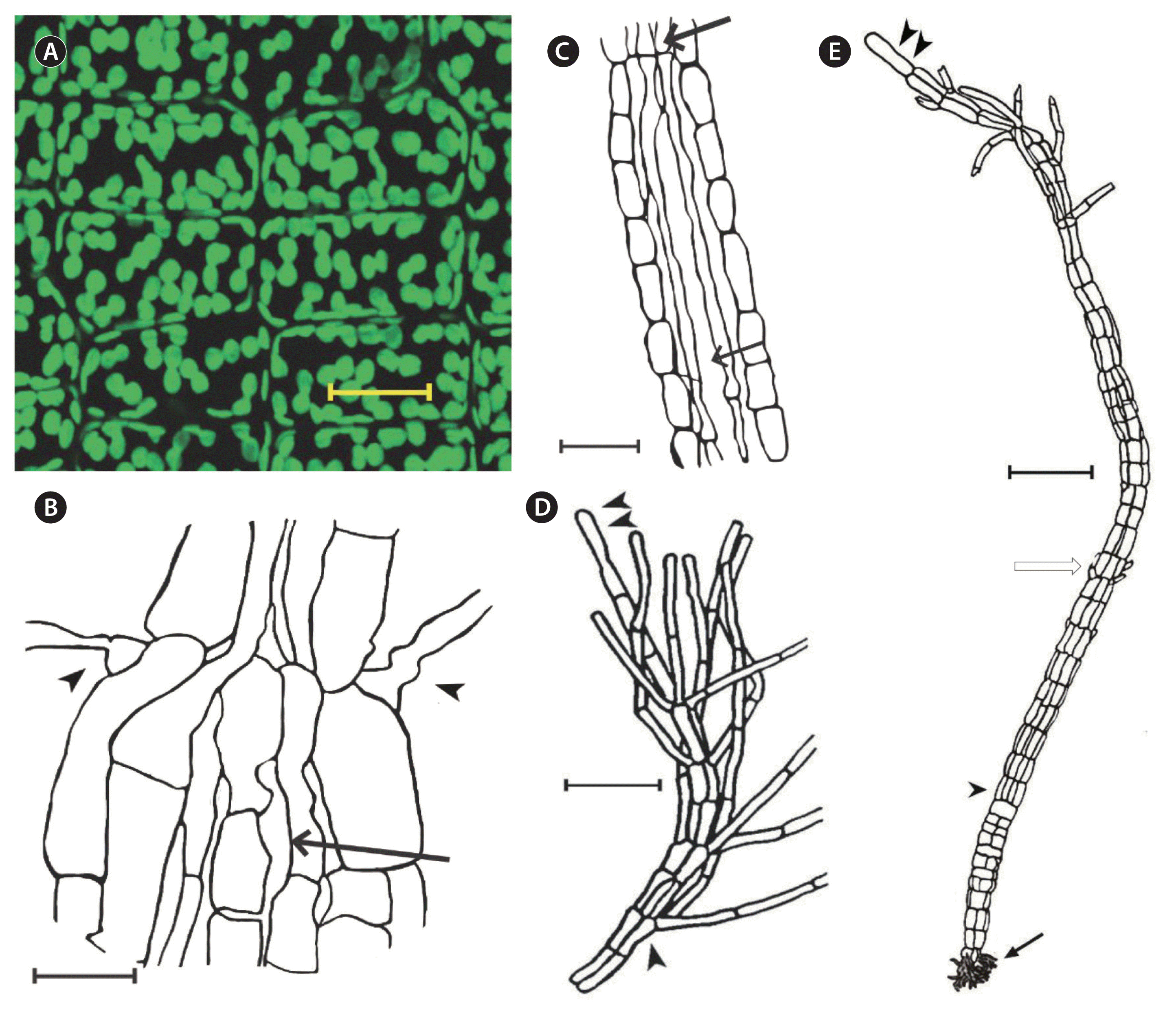
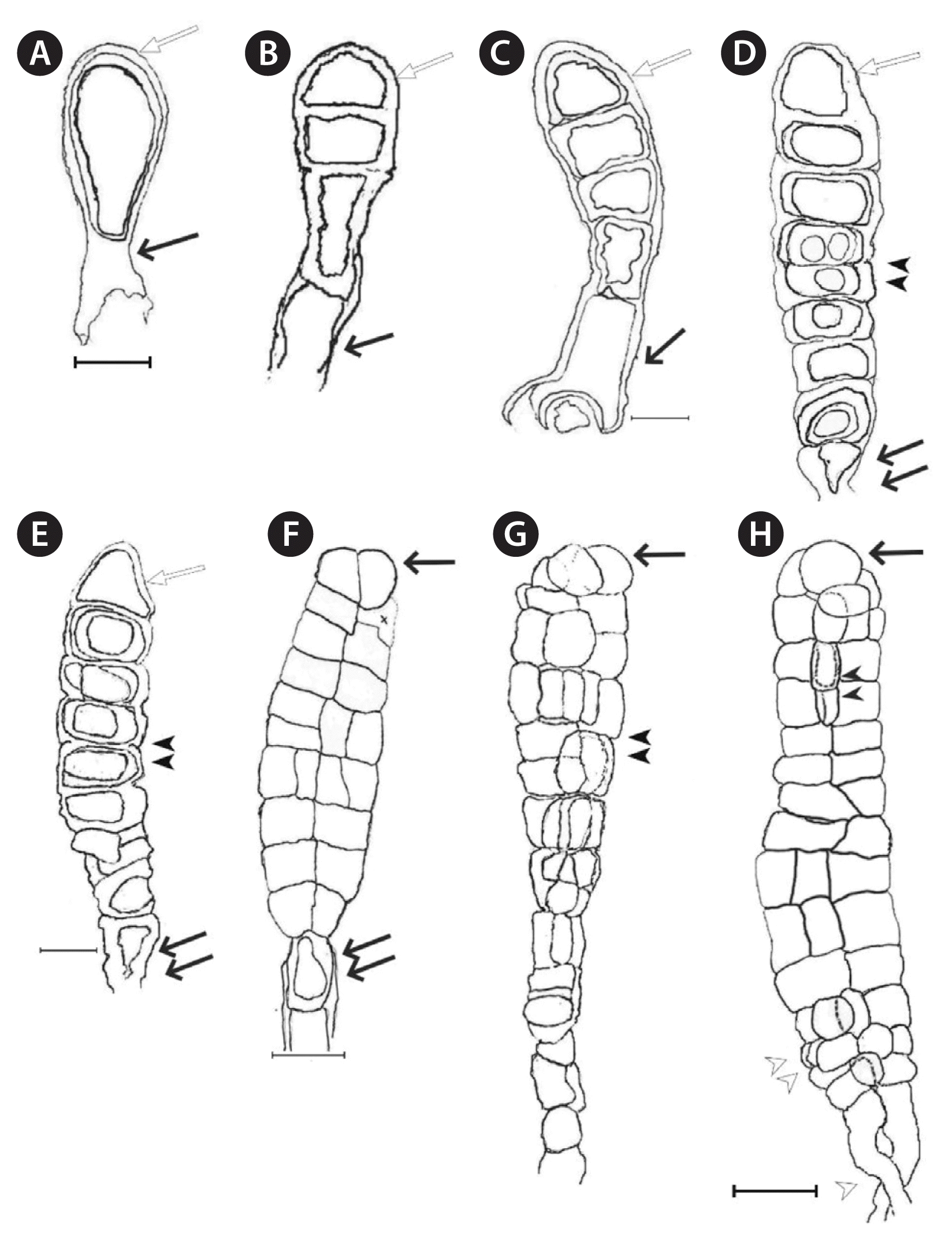
Platysiphon verticillatus. (A) Cortical cells in uniform arrangement with non-pyrenoidal plastids from median, expanded thallus. (B) Origin of photosynthetic filaments in upper rat-tail (arrowheads); note lengthy medullary cells (arrow); slide no. 11565. (C) Medullary filaments in upper rat-tail (arrows); note their bulbous linear connections, considerable length and occasional lateral fusion slide no. 11716. (D) Apical cell (double arrowhead) and pigmented filaments (arrowhead) in upper portion of juvenile specimen; fragment of type collection. (E) Juvenile specimen with terminal apical cell (double arrowhead), attachment cells (arrow), early stages of parenchyma in the stipe region (arrowhead), first cells (white arrow) of pigmented filaments specimen from type collection; slide no. 11567. Scale bars represent: A, 20 μm; B & D, 10 μm; C & E, 25 μm.
Platysiphon verticillatus. (A) Cortical cells with regular arrangement from median thallus, empty, discharged small (arrow), and large (arrowhead) cysts on the thallus surface. (B) Photosynthetic filaments from cortical cells on expanded thallus. A & B, scanning electron microscope (SEM) images. (C) Hydrated protoplasts, first reproductive stage (arrows), note the plurality of discoid plastids in adjacent cells. (D–F) Protoplast divisions: equal (D & E, arrows), unequal (E, protoplasts & F, empty cysts size, double arrow). (E) Note the morphology change of the upper and lower protoplasts (white arrows), protoplast attachment to cell wall (arrowhead); note evidence of an incomplete cell wall between protoplasts (double white arrow), completed wall pore (white arrowheads). (F) Reproductive thallus with large protoplast attached to the inner face of the cell wall in early stage of wall pore development (double arrowhead), and adjacent small protoplast (arrowhead) with wall pore (arrow) completed, cell wall separating small and larger protoplasts (white arrow), pore beneath empty small protoplast (arrowhead); absence of parenchyma in the reproductively mature thallus, and also the difference in the discharged cysts size (joined arrows) and the presence of a pore beneath the smaller, empty cyst (double arrow). (G) Reproductive thallus in later stage of pore and cyst formation; note the membrane residue (double arrows) remaining in the parent cells of the larger and smaller protoplasts; note also, the cell wall (arrow) separating the two parent cell, also the difference in cyst diameter (joined arrows), discharge pore in empty cyst (arrowhead), and the absence of parenchyma. (H) Multiple, replete (arrow) and discharged cysts (joined arrows) on a reproductively mature thallus surface; SEM stub number: 040806. Scale bars represent: A & C, 10 μm; B, 100 μm; D & E, 20 μm; F–H, 25 μm.
There is little value in the repression of long-standing previous biological results as systematic criteria (Nilsson et al. 2006, Augyte et al. 2018, Ledford 2018). Gross and cryptic biological features compliment use and value of genetic protocol (Lane et al. 2006, 2007, Nilsson et al. 2006, Mann 2010, Hu et al. 2012, Leliaert et al. 2014, Campagna et al. 2017, Díaz-Tapia et al. 2017, Leliaert and De Clerck 2017, Páll-Gergely 2017). We recognize similarity of core Halosiphon Clade genes, especially in Pl. verticillatus and Pu. glacialis. Molecular criteria are undeniably successful to identify the generic relationship, including Halosiphon tomentosus, Stschapovia flagellaris, Pl. verticillatus, and Pu. glacialis, and their inclusion in the Stschapoviales (Kawai et al. 2015a). We submit that products of evolution, minute and gross biological differences, result from a variety of genetic reshuffling, different in each taxon. Conspecificity based on several shared, identical genes, is questionable.
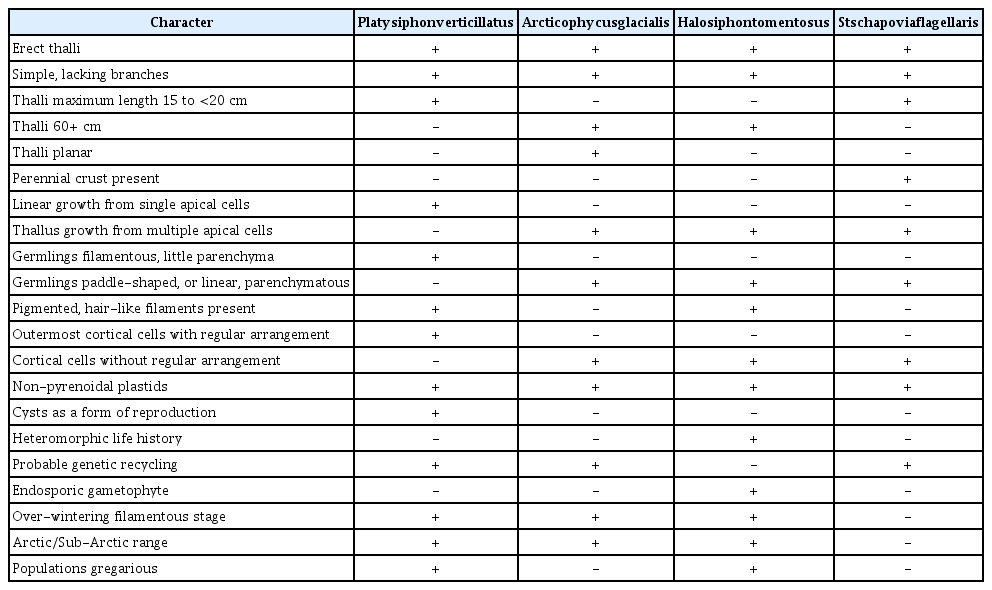
We identify and strengthen the value of the pooled use of phylogenetic and morphological data to identify taxa (Table 1) (Páll-Gergely 2017). In support of this premise, we identify and attempt to clarify all the known biological features of Pl. verticillatus and Pu. glacialis. We document unidentified, novel, reproductive stages in both species, and the previously unknown early distinctive growth phases of Pu. glacialis. We posit the life history of both species, suggest necessary means of over-wintering and cite their known distributions. The new genus Arcticophycus is described; Platysiphon, Platysiphonaceae and Stschapoviales are emended. A. (Arcticophycus), Pl. (Platysiphon), and Pu. (Punctaria), abbreviations to be used hereafter.
Comparisons of morphological characteristics of four species
The binomial Punctaria glacialis is unacceptable. It differs from Platysiphon and Punctaria at the molecular level and by many other biological differences including the presence of pyrenoidal plastids and its different reproductive strategies (Fletcher 1987, Wilce and Pedersen 2003, Wilce and Bradley 2007, Kawai et al. 2015b).
MATERIALS AND METHODS
We use all the data from specimens of Pl. verticillatus and Pu. glacialis collected in situ in Ragged Channel, N Baffin Island during three light seasons, and from NW Greenland (Fig. 1D) from subsequent Amherst laboratory studies (1994–1998), our own, and those of Kawai et al. (2015a), and importantly, the literature, correspondence, and historical specimens of Pu. glacialis from NW Alaska and the Canadian Archipelago (Lee 1980). Late July/August 1994 collections of inshore sediments contained A. glacialis germlings and reproductively mature thalli, both significant in our understanding of the life history and development of Pu. glacialis. Pu. glacialis which has been seen in situ by Lee (1973–1975) and Wilce. Those who collected and studied Pl. verticillatus in situ are Saunders, Küpper, Kawai, and Wilce.
Specimens of Pl. verticillatus and Pu. glacialis thalli from Ragged Channel prepared for electron microscopy were fixed in 2.5% glutaraldehyde and post-fixed in 1% osmium tetroxide. They were dehydrated with an ethanol series and critical point dried with CO2. The material was coated with gold with a sputter coater and viewed with a JSM5600LV-scanning electron microscope (JEOL, Tokyo, Japan). Details concerning electron microscopy protocol are in Chandler and Roberson (2009) and Hayat (1981). A Wild M20 microscope (Wild Instruments, Heerbrugg, Switzerland) was used for all light microscopy; the Wild Camera Lucida and drawing arm was used to make line illustrations.
RESULTS
Platysiphon verticillatus Wilce 1962; Wilce & Bradley, emended
Type specimen
A juvenile specimen collected Aug 23, 1960 near the old village site of Qaanaaq, NW Greenland; slide no. 11731; MASS.
Synonymy
Platysiphon gracilis Kawai et al. 2015a.
Distribution
The old village site of Qaanaaq, Inglefield Bay, Thule District, NW Greenland (Fig. 1D, arrow 1); mature and juvenile thalli were found throughout the summer light season in Ragged Channel, N Baffin Island, Nunavut, Canada (Fig. 1D, arrow 2). One deep-water site, Sep 2015, N Labrador, Canada; ca. Nome, AK, both collections by G. W. Saunders (personal communication). Data not shown in Fig. 1D. Ragged Channel populations are positively annual in occurrence.
Vegetative morphology
Thalli of Pl. verticillatus are initially filamentous, later tubular and flaccid with modest lateral expansion (Fig. 1A & B) (Wilce 1962, fig. 1). Thallus length primarily from an apical cell (Fig. 2D & E). Cortical cell shape is variable, rectangular, some ovoid to globose 8–12 μm diameter, lacking all differentiation (Figs 2A, 3A & B). Derivatives of the apical cell divide periclinally to form a weak medullary layer of one cell thickness, these cells become extremely long, bulbous at cell junctions (35 to 65 μm) (Fig. 2B & C). Cortical cell arrangement in vertical and lateral rows, packets of cells evident, reflects orderly cell divisions (Figs 2A, 3A & B). Pigmented filaments (8–12 μm wide, ≥60 μm long) from cortical cells, mostly in tight clusters, often whorled in arrangement in the cells are numerous in median and distal portions of the thallus (Figs 2D & 3B) (Küpper et al. 2016, fig. 3b & c). Medullar cells have long rib-like appearance at the cortex inner surface. Thallus weakly parenchymatous basipetally, merges into a tubular lower blade (Fig. 1A). Holdfast is initially several short tubular cells, with thallus maturity a small pad; stipe short (1–3 cm) (Fig. 2). Numerous non-pyrenoidal plastids cover the exterior face of each cortical cell (Fig. 2A). Mature thalli consist of a solid stipe, a disc holdfast, and a completely hollow, expanded, tubular mid-portion, lacking appreciable parenchyma except in proximal and distal regions. The distal portion of the thallus (the “Rat Tail”) (Fig. 1A & B) (Wilce 1962, fig. 1, Küpper et al. 2016, fig. 3b & c) is distinctive for this taxon, generally one-third in length to the expanded region, becomes heavily invested with lateral pigmented filaments and invariably with the epiphyte Phaeostroma pustulosum Kuckuck (1893).
Reproductive morphology
All Ragged Channel mature Pl. verticillatus thalli remain vegetative through the middle of September. Observations of viable in situ thalli and laboratory experiments suggest reproduction is initiated under low incident light after a period of semi or total darkness. Late autumn intermittent low light permits development of over-wintering filamentous reproductive stages. Possible vernal reproduction is suggested by studies of thallus fragments collected in early September and cultured in our laboratory. In this scenario, survival of over-wintering filaments or thallus fragments must be present to initiate reproduction and germling development. In either instance we posit the following developmental stages leading to reproductive propagules. Prior to incipient reproductive stages plastids and other cytological organelles cover the full face of all cortical cells (Fig. 2A). Cytological change in cortical cells occurs with severe cellular hydrolysis resulting in compressed cell contents within the spherical protoplast (Fig. 3C). One of three early reproductive stages follow: equal protoplast division (Fig. 3D, arrow), unequal protoplast division (Fig. 3E, double arrow) or as in the greater majority of cells, direct morphology change of the protoplast without division. Whether an equal or unequal division, a cell wall forms to separate the two hydrated protoplasts (Fig. 3C, arrows). The only function attributed to protoplast division is a substantial increase in subsequent cyst numbers accompanied by uniformly smaller (<2/3) diameter cysts following a previous protoplast division. In each instance of early reproduction, the distal end of a slightly elongated protoplast becomes attached to the inner face of the parental cell wall (Fig. 3F, double arrowhead). A chemical dissolution of the wall occurs with the formation of a pore (Fig. 3E, arrowhead, F, arrow). The naked protoplast emerges, becomes spherical, forms an investing membrane, followed by a thin cyst-like wall, similar in shape to the cyst morphology of many Chrysophytes (Olli 1996). Unidentified cellular materials, including the cellular membrane of the attached protoplast, remain in the vacated cell (Fig. 3G, double arrows). Cysts vary in size (6.5–12.0 μm) depending on proportions of previous protoplast divisions. Undivided protoplasts form the greater number of cysts. Whether small cysts function to form filaments, as we believe occurs with the larger cysts remains unknown. Under laboratory conditions a myriad of globose, densely pigmented cysts was liberated from parental cells. Replete (Fig. 3H, arrow) and discharged (Fig. 3H, joined arrows) are distinctive and numerous on thallus surface (Fig. 3H, arrow). Cysts walls are smooth, unornamented, with only a delicate reticulate pattern (Fig. 4A & B). A biflagellate motile cell is released from each cyst (Fig. 4C). Early stages of subsequent filamentous growth with hyaline unsheathed hairs (Fig. 4D & E, arrows) occurred under laboratory conditions. Zoospores were liberated from sporangia in culture (Fig. 4F & G, double white arrow). Comparable in situ filaments with sporangia that liberate zoospores are likely responsible for under-ice germling development in spring (May–June, Ragged Channel site). Ragged Channel populations are positively annual in occurrence.
Platysiphon verticillatus. Mature cysts, zooid discharge and subsequent reproductive filament development. (A) Replete cyst; note the delicate irregular pattern on the cyst wall and irregular cyst collar (arrow); scanning electron microscope (SEM) stub number: 040806. (B) Discharged cyst; note the uneven rupture of the discharge pore; SEM number: 040806. (C) Zooid (arrow) released from cyst (double arrow); note the increase in zooid size in contrast to cyst diameter. (D) Branched filaments (arrows) in culture derived from a germinated cyst zoospore and the irregular knobby cells (arrowhead) (pre-sporangia?), presence of hyaline non-sheathed hairs (arrows), branched, knobby filaments (double arrow). (E) Portion of a hair and its origin (arrow). (F) Filament with terminal (white arrow) and lateral sporangia (double arrow), note zooids (double white arrow) released from an intercalary cell (double arrow); note the presumed early sporangial development in numerous irregular globose cells (arrowheads), sporangial cells (arrows). (G) Terminal sporangium replete with spores (arrow) and an intercalary sporangium (arrowhead) with discharged spores (double white arrow). Scale bars represent: A & B, 4 μm; C, 20 μm; D, 50 μm; E & G, 10 μm; F, 30 μm.
Arcticophycus gen. nov. R. T. Wilce & P. M. Bradley
Genus description
Erect thalli have undivided blades up to 50 cm in length (Fig. 1C) (Rosenvinge 1910, fig. 6, type), parenchymatous throughout; germlings initially attached by zooid cell wall (Fig. 5A–C, arrow & 5D–F, double arrow) later by non-photosynthetic, elongate tubular cells (Figs 5H, white arrowhead & 6A–D, arrow), followed by a definite pad (Fig. 6D, white arrow). Cortical cells variable size, arrangement, morphology and cell wall thickenings, phaeophycean hairs and photosynthetic assimilatory filaments are absent. Plastids lack pyrenoids (Fig. 7C). Reproduction occurs with the formation of thick walled sporangia (Type 1) (Figs 7F, arrows, 8H, double arrow, 8J, arrow & 8K, white arrow) and also from cortical cell derived sporangia (Type II) (Fig. 9A, arrow).
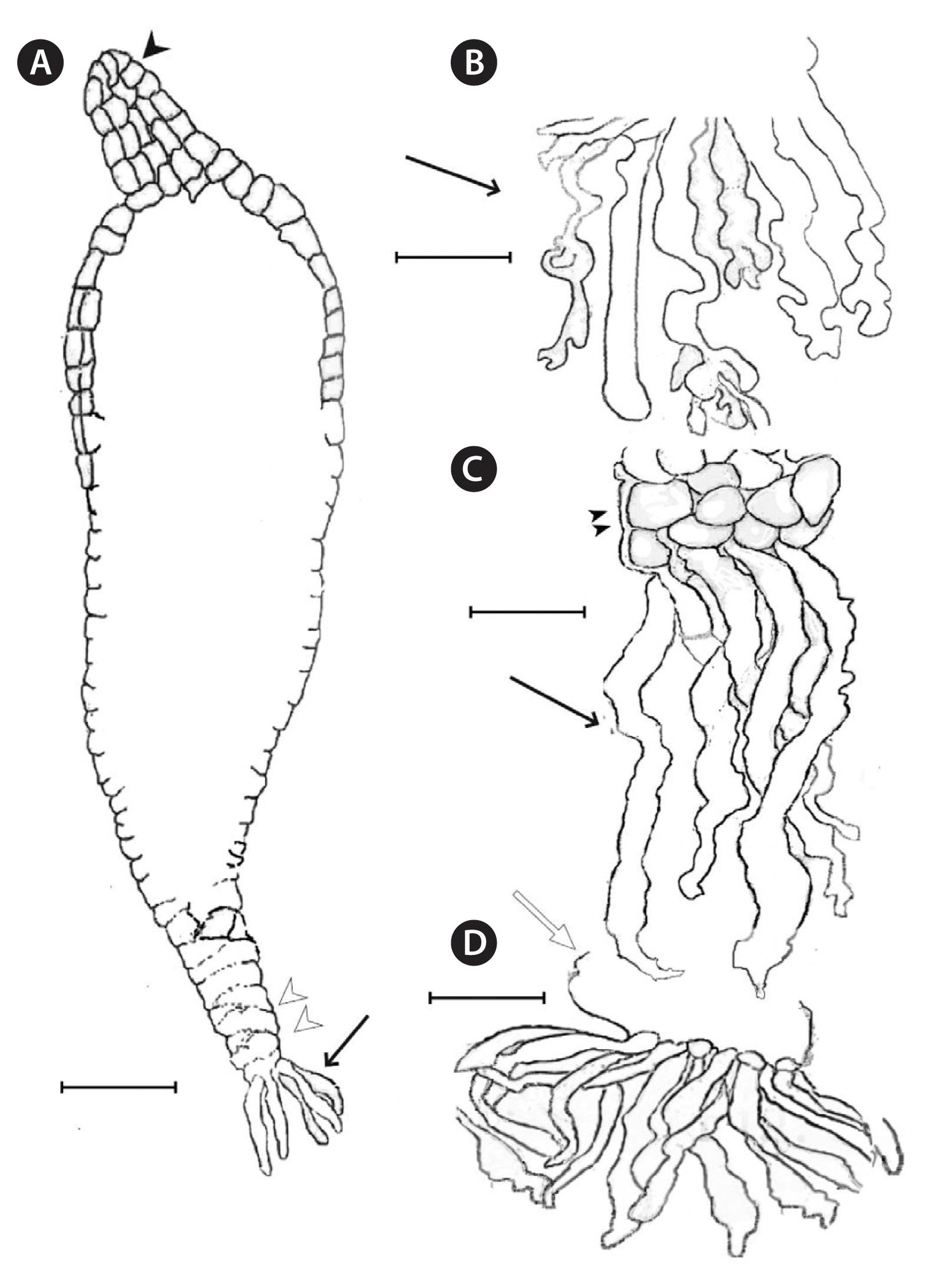
Arcticophycus glacialis. Mature germling with parenchyma in the stipe region and germling attachment structures. (A) Germling (242 μm long, 32.96 μm broad); note the presence of single cell attachment tubular cells (arrow); note also, multiple apical cells (arrowhead), absence of attachment disc initiation and the development of stipe length (double white arrowhead); slide #11180. (B–D) Lower portions of germlings with well-developed tubular cells and highly irregular cell wall edges (arrows). (C) Initial cellular development of the attachment disc (double arrowhead) formed immediately above the unicellular, tubular cells. (D) Later stage in disc initiation showing parenchymatous bulge (white arrow); slides #11180–81. Scale bars represent: A, 50 μm; B & C, 35 μm; D, 65 μm.
Arcticophycus glacialis. Filament to parenchymatous development and modes of early attachment, specimens from moderately exposed, 2–3 m depth, Ragged Channel, Baffin Island Sep 1994, R.T.W. Slides no’s 11180–11182. (A) First cell (white arrow) of erect germling and the remains of zooid attachment cell wall (arrow). (B & C) Filamentous germling stages: zooid unidirectional germination, remains void after cellular division to form the first filament cell (B–E, white arrows); note the apparent role of the zooid wall in its early attachment (A–C, arrows). (D–F) Germlings with first formed parenchyma (double arrowhead); note the undivided apical cells in (D & E, white arrows) and the distinctive triangular morphology of the first formed pigmented cell (D & F, double arrow). (F) Germling (148 μm long), parenchymatous with multiple apical cells (F–H, arrows); note the characteristic shape of the first filament cell and its relation to the initial attachment cells (E & F, double arrows). (G & H) Post-filamentous germling with first formed parenchyma (double arrowheads) in upper germling and basal parenchyma (H, double white arrowhead) that initiate disc holdfast; note multiple apical cells (arrows), and first formed attachment tubular cells (white arrowhead); slide no. 11182. Scale bars represent: A, 20 μm; B–F, 10 μm; G & H, 25 μm.

Arcticophycus glacialis. Mature linear germling, historic Punctaria stipitata collections, gross cytology, mature reproductive specimens and species reproductive stages, sensu Rosenvinge (1910) (type specimen). (A) Linear, late stage germling attached to lose lying, elongate thalli at ca. 8–10 m from a little disturbed environment. Note multiple apical cells (white arrow), and parenchyma throughout (arrowhead). Note also, the absence of the mature attachment disc initiation (arrow) (slide no. 11180). (B) Collection by F. Johansen (arrows), Port Clarence embayment, AK, Jul 13, 1918, identified as Punctaria stipitata Collins, portion of type collection; note stipe and holdfast (white arrows). (C) Surface of cortical cells with non-pyrenoidal plastids. (D) Multiple clusters and frequently confluent pear-shaped sporangia; Ragged Channel, Baffin Island, R.T.W. (E) Reproductive and partially vegetative mature specimens (arrowhead), Ragged Channel, N Baffin Island, ca. 8–10 m. Sep 1994, R.T.W.; note the extensive loss of thallus (double arrowhead) following sporangium discharge and elongate stipes, discoidal holdfasts (arrows). (F) Reproductive drawings form the type specimen (Rosenvinge 1910, p. 120); note presence of parenchyma, pear-shaped sporangia (Type I) (arrows) and irregularity of cortical cell size and arrangement. Scale bars represent: A, 20 μm; B, 3 cm; C, 25 μm; D, 50 μm; E, 8 cm; F, 340×. [Colour figure can be viewed at http://www.e-algae.org].

Arcticophycus glacialis. Stages in the development of zooids and branched filaments taken from specimens attached to lose lying thalli at ca. 8–10 m. Asterisks identify dividing cells. (A & B) First divisions of sporangial thick-walled cells (arrows). (C & D) Early developmental stages of thick walled to form either zooids (C, double arrow); note nascent filament cell with a single plastid (C, arrow), discharged thick-wall cell (C, arrowhead) or filaments (D, double white arrowhead), discharged thick-wall cell (D, arrowhead). (E–H) Aggregates of thick-walled cells within sporangia husk (arrows); note also, germination of thick-walled cells within sporangium to form the filaments (E & F, double white arrowhead) and the presence of one belt-shaped, non-pyrenoidal plastid in each filament cell (arrows). (G) Filament development from thick-walled cell showing first anticlinal apical cell division (white arrowhead) in the early development of a branched filament; note also, the larger proportion of discharged cells (H, arrowheads) to those that initiate branched filaments (F–H, double white arrowhead). (H) Later stage in the role of sporangial thick-walled cells attached to parental thalli. Note cell with three protoplasts (double arrow, presumably zoospores) and numerous adjacent discharged cells (arrowheads); note also, filament initial development (double white arrowhead). (I) Thick-walled sporangium cells; note their cell walls (arrow) and sporangium wall (double arrow). (J) One thick-walled cell within the sporangium wall (arrow), cell wall (double arrow). (K) Ruptured thick wall sporangial cell (white arrow) with retained protoplast (arrow). (L–N) Scanning electron microscope image: zooid discharge (white arrows), remnants of sporangium wall (arrows). Branched filaments (arrows) within remnants of sporangium wall (white arrows), from herbarium specimens no.’s 00387893-94 Ragged Channel, Baffin Island ca. 8–10 m. Scale bars represent: A–D, G, H & J, 20 μm; E, F, I & K–M, 10 μm; N, 50 μm.

Arcticophycus glacialis. Sporangium (Type II) and early filament development. (A) Thin-walled, rectangular sporangium (arrow) with thin-walled spores (double arrow), note also non-pyrenoidal plastids in adjacent cells (double white arrow). (B & C) Hylain zooid cover (double arrowhead), germ tube continuous with zooid cover (arrow), single cup-shaped plastid (arrowhead). (C) Second filament cell (arrow), of presumed over-wintering filament, as opposed to germling development (see Fig. 5A–D). A–C from reproductive parent thallus. Scale bars represent: A, 10 μm; B & C, 25 μm.
Arcticophycus glacialis (Rosenv.) R. T. Wilce & P. M. Bradley, emended
Type
Rosenvinge (1910, p. 118, fig. 6), “Floating in the ice, along the E side of Store Koldewey Island, 7–8 km SSW of Danmarks Havn, NE Greenland; Aug 26, 1906, leg., Andr. Lundager, no. 831, COP. (! = R.T.W., I have seen); reproductive with unilocular sporangia.” This largely complete specimen (lacks the holdfast and a small (?) portion from the blade tip). A blade fragment (14–15 cm long, vegetative) from this collection is present in the NYBG herbarium, no. 721 (!) but the date cited incorrectly as Aug 28, 1907, leg., Andr. Lundager.
Synonymy
Punctaria glacialis Rosenvinge 1910, Vol. 43, pp. 93–133, figs 6 & 7.
Punctaria fissilis Setchell & Gardner 1924, Vol. 8(III), p. 4; 1925, p. 518.
Punctaria stipitata Collins 1927, Pl. 1, figs 8 & 9.
Platysiphon gracilis (Rosenv.) Kawai et al. 2015a.
Species description
As for the genus, see above.
Morphology
Thalli from Ragged Channel collections (Fig. 1B & C), develop from a stipe 2.5 cm long, broadly oblong to narrow linear, more Saccharina-like than the leaf shape of a plantain, i.e., the type specimen, largest, and distally incomplete, 66 cm long, 5.75 cm broad (Kawai et al. 2015b, fig. 4a). Holdfasts of germlings are composed of a cluster of long tubular, thin wall hyaline cells, with irregular margins (Fig. 6A–D, arrows). A disc holdfast forms immediately above the germling holdfast (Fig. 6C, double arrowhead). Blades are entire without ruffled margins. Mature specimens are fragile, despite apparent coarseness, fragmenting readily in situ when handled, even more so as dried herbarium material. As an epiphyte of sublittoral Fucus thalli resemble juvenile Saccharina latissima or Petalonia fascia. In situ specimens range in color from a tannish brown to deep brown.
Reproduction
A. glacialis forms two sporangial types (Types I–II), distinguished by their mode of occurrence, morphology, wall thickness, size, and their ultimate zooid development. A. glacialis sporangia Type I contains thick walled cells that release zooids and branched upright filaments. Zooids released from A. glacialis sporangia (Type I, see Rosenvinge 1910, fig. 7e) give rise to annual germling populations (Figs 5A–H, 6D & 7A). Zooids attached to substrata divide to form the first pigmented cell of a short filament (Fig. 5B & C, arrows). The zooid cell wall functions as the initial attachment structure (Fig. 5A–D, arrows). Later attachment occurs by ephemeral, hyaline, elongate tubular cells with irregular margins that show no evidence of further cell division (Figs 5H & 6A–D, arrows). Fletcher et al. (1984) and Fletcher (1987, fig. II, 18) illustrates these structures as “rhizoidal filaments” formed from the base of a Sargassum muticum germling and, variously, with other taxa as: “fibrous holdfast, unicellular multicellular filaments, or a fibrous mat of rhizoidal filaments.” These attachment structures are neither rhizoidal or filamentous, rather elongate, saccate, non-pigmented cells that putatively contain an adhesive material, released to enhance juvenile thallus attachment (Baker and Evans 1973, Toth 1976, Fletcher 1980). The initial two tubular cells develop from a triangular shaped first cell of the filamentous germling (Fig. 5D–F). Subsequent attachment cells develop from the basal stipe region (Figs 5H, double white arrowhead, 6B & D, arrows), lost with the development of the disc holdfast of mature thalli. Disc holdfasts are initiated in the swollen, bulbous parenchymatous region immediately above the juvenile attachment structures (Figs 5H, double white arrowhead & 6C, double arrowhead). Thallus parenchyma is initiated early in A. glacialis juvenile development (Fig. 5D–H, double arrowheads). Germlings soon expand laterally to form a single layered, one to several mm long paddle-shaped juvenile blade (Fig. 6A). Peripheral blade regions ultimately become thickened from the median thallus toward its periphery in an irregular jagged, saw tooth manner (unlike the uniform lateral blade development in Saccharina latissima). Mature thalli are solid throughout, the median axis with 4–8 cortical and medullary cell layers, 45–60 μm thick (Rosenvinge 1910, fig. 7b–e, Kawai et al. 2015b, fig. 1a–l). The cortical cell layer of A. glacialis is distinctive by its outer irregular cells in palisade-like arrangement. Cortical cells in surface view measure 8–12 μm. Medullary cells are ovoid, never elongate, and measure 9–15 μm diameter.
Specimens of A. glacialis collected at 2–4 m in late July/August and September contained soral reproductive areas (Fig. 7D). Their appearance in A. glacialis is similar to the contiguous soral areas in foliose Ulva blades (Fig. 7D). Several to many thick-walled cells formed in each sporangium (Type I) (Figs 7F, arrows, 8J, arrow & 8K, white arrow), all highly pyriform with greatly swollen basal regions and a neck-like distal region (Fig. 7F, arrows). Their diameter is similar to the diameter of an undifferentiated cortical cell. These cells divide further to form a multi-cell population of thick-walled cells enclosed by the sporangium wall. The following comment of Rosenvinge 1910 (loc. cit.) shows the reproductive features he recognized and also his reluctance to fully sanction the character of what he describes as “unilocular sporangia.” Viz., “Strange to say, the zoospores had not developed normally but had formed cell walls (Figs 7F & 8A–H) without having been set free, and the older sporangia thus became closely packed with polygonal cells, which gradually became rather poor in contents and might sometimes suggest the structure of a plurilocular sporangium” (Fig. 7F, arrow). The ruptured A. glacialis sporangium wall becomes a surrounding husk (Fig. 8F, arrow & J–N, arrows) to contain numerous protoplasts. Each protoplast becomes thick walled, ultimately transformed into a 4-chambered plurilocular sporangium their contents released as zoospores (Fig. 8F–H, arrows), or to germinate in situ to form branched filaments (Fig. 8H, double white arrowhead, M & N, white arrows).
A. glacialis sporangium Type II occurs sporadically, not aggregated on the thallus surface, rectangular, larger than Type I sporangium, regular cell wall thickness (arrow), contain only pre-zooid cells, zooids larger than those in Type I sporangium (Fig. 9A, double arrow). Zooids germinate to form filaments (Fig. 9B & C). We posit that these filaments are the over wintering portions of the species vegetative life history, to give rise to vernal zooids that develop erect A. glacialis thalli. It is likely that A. glacialis over-wintering strategy is similar to that of Pl. verticillatus.
Occurrence
A. glacialis is found throughout NE and NW Greenland and the N American Arctic (Fig. 1, ellipses): Canadian Archipelago: Lee 1974–1975; Wilce, Devon Island, Aug 1963, Grise Fjord, Ellesmere Island, Sep 1969, Ragged Channel, N Baffin Island, Nunavut, Canada 1979, 1981, 1983, 1994; Saunders, Ragged Channel, 2014; NW-NE Greenland: Rosenvinge 1910;Lund 1933 (!), 1959a, 1959b (2 locations); Wilce 1962, ca. Qaanaaq, Inglefield Bay; Mestersvig (one specimen, no collector identified (!); N-NW Alaska: Dube, Aug 1978, “attached to small rocks in shallow water; juvenile and vegetative, vicinity of Point Barrow and Cape Lisbourne” (!); Johansen, Jul 30, 1913, Port Clarence, dredged in 6–9 m, station 20 b–c (Fig. 1B & D, arrow 2) (!); National Herbarium Canada, Ottawa no. 4255, University of California Herbarium no. 207083 (!); Dunton, Kaktovik Lagoon, ca. Prudhoe Bay, Beaufort Sea, Aug 21, 2012, no. 9119 (Fig. 1D, arrows 3) (!); Nome, AK, Saunders.
Notes on the initial Punctaria glacialis NW Alaskan collections
Details of the history of A. glacialis concerning the publication of this species are interesting, some unfortunate and unforeseen. F. Johansen, a remarkably intrepid collector of marine species, a member of the Canadian Arctic Expedition, 1913–1916, made collections of Pu. glacialis in NW Alaska, Jul 30, 1913. These collections included three complete juvenile specimens (Fig. 7B) of a thalloid brown alga, which were ultimately sent to F. S. Collins in 1916 for identification. On Feb 26, 1917, Collins wrote to W. A. Setchell for an opinion on the three specimens that he included with his letter, which “are more like Punctaria than Laminaria” and requested Setchell, “Please return the specimens.” Collins wrote to Setchell 18 months later on Aug 4, 1918 and again asked for Setchell’s comments on the “unknown Punctaria.” Setchell promptly replied, Aug 13, 1918, writing that he found “hairs and what may possibly be unilocular sporangia on your plant. It can’t be Pu. glacialis Rosenvinge (1910), if his description is correct, i.e., Rosenvinge. A new species?” The three specimens Collins sent to Setchell remained in the herbarium of the University of California, Berkeley, ultimately to become type material of Punctaria fissilis Setchell & Gardner (1924, p. 4). In late 1918 or early 1919, Collins completed his manuscript on the marine algae of the Canadian Arctic Expedition including his description of Pu. stipitata sp. nov. Collins died May 25, 1920, before the manuscript was submitted for publication. Four years later Setchell & Gardner published Pu. fissilis (1924), probably unaware of Collins unpublished description of Pu. stipitata. The Collins algal collections and manuscripts were ultimately sent to the New York Botanical Garden, at the time directed by M. A. Howe. Howe, unaware of the Setchell and Gardner 1924 publication, subsequently published the Collins report without change, Nov 24, 1927, hence the lack of priority for Pu. stipitata Collins. Howe retained a single incomplete thallus of the species and returned four specimens and a small blade fragment to the Canadian National Herbarium with Howe’s annotation “part of type,” accompanying each specimen. Collins incorrectly writes on his label that the collections were made in 1910 as opposed to 1913 and adds that the “bottom: [was] mud and sand, at 2–3 fathoms.” The entire Johansen collection consists of eight thalli and a few disassociated blade fragments. Four of these specimens are complete with holdfasts, stipes and most of their blades, all now understood as juvenile. Seven thalli show similar morphological features, all of which have undivided blades. One of three specimens in the California collection has a blade divided several times in narrow laciniate segments, this being the rational for Pu. fissilis. No evidence of hairs or photosynthetic filaments or reproduction is present on these juvenile specimens (!).
Thus, the status of Pu. glacialis remained unchanged. A new understanding of the taxon and it relation to Pl. verticillatus takes into account recent information from the following: 1) the numerous collections of the species by Lund in E Greenland, specimens from Lee and his divers in the central Canadian Archipelago, studies of Wilce in NW Greenland and the Canadian archipelago; 2) the laboratory cultures of Pl. verticillatus in Amherst; 3) the electron microscope studies of Bradley, and the molecular studies of Ragged Channel specimens collected by Wilce and Saunders, and those provided to Kawai; and 4) several related publications (e.g., Rosenvinge 1910, Pedersen 2011, Kawai et al. 2015a, 2015b, Mathieson and Dawes 2017).
DISCUSSION
The relationship of Platysiphon verticillatus and Arcticophycus glacialis, indeed their identities, has been troublesome. A. glacialis, enigmatic from its discovery, became more so with the recognition of Pl. verticillatus. Despite the current general acceptance of molecular criteria to establish genera and species, we describe Arcticophycus as novel based on taxonomic priority and a multitude of its distinct biological features. Our separation of Pl. verticillatus and A. glacialis is also based on biological characteristics (Table 1) viz.: small thalli (max. 15 cm) vs. large thalli (>50 cm), delicate, and largely tubular, with numerous photosynthetic filaments vs. coarse, solid parenchymatous, devoid of hairs or photosynthetic filaments. Contrast between species is also evident in their mode of growth. A single apical cell in Pl. verticillatus is responsible for linear growth. Multiple apical cells of A. glacialis form soon after a short filamentous stage (Figs 5F–H & 6A), these to produce juvenile paddle-shaped germlings by repeated periclinal divisions (Fig. 6A), and subsequent linear growth. Medullary tissue development (Wilce 1962, figs 1–15) differs between the two species: Pl. verticillatus has a few medullary cells, initially globose, fail to divide but become extremely elongate, bulbous at their junction and frequently laterally connected (Fig. 6B & C, arrows). This contrasts with the parenchymatous structure and uniform globose to ovoid medullary cells in A. glacialis (Rosenvinge 1910, fig. 7b–e, Kawai et al. 2015b, fig. 1a–l). Outer cortex cells of both species show contrast in cell arrangement, plastid size and number. Plastids of Pl. verticillatus (2.3–5.0 μm) (Fig. 2A) are slightly smaller than those of A. glacialis (3.0–4.0 μm) (Fig. 7C).
Pl. verticillatus populations at the Ragged Channel site (2–4 m) are locally expansive, strongly gregarious. Thalli of A. glacialis collected from Baffin, Devon, and Ellesmere Island occur sporadically, attached immediately beneath the sublittoral barren zone (Wilce 2016), ca. 2–7 m below low water level, epiphytic on Fucus (Lee 1980), never as gregarious populations but singly, or in a weak cluster of individuals. This occurrence is unlike that described by Lund (1959a, p. 135) “… the species at depth from 4–38 m in E Greenland from collections made with a Petersen Grab Lund.” Based on in situ observations (R. T. Wilce) of the species, the deeper collections cited by Lund were likely unattached, loose-lying thalli.
Known reproduction in Pl. verticillatus and A. glacialis differ. Occurrence of a novel brown algal cyst, in Pl. verticillatus (Wilce and Bradley 2007, Wilce 2016) is unique. Zooids liberated from cysts, form filaments, filaments with sporangia in laboratory culture, suggested zoospores to germlings in situ. We posit an asexual life history for Pl. verticillatus accompanied by genetic recycling. Reproduction in A. glacialis occurs by zooid release from sporangia (Type I) to form germlings (Figs 5A & 6A). Also, from a second form of sporangia, sporangia (Type II) (Fig. 9A). We posit that zooids released from these sporangia form filaments (Fig. 9B & C), filaments responsible for species survival through the winter dark season. We also suggest that vernal germlings develop from zoospores discharged from sporangia formed on the over-wintering filaments, followed by a new generation of erect thalli. Similar to Pl. verticillatus, A. glacialis appears to have an asexual life history with recycled genetic constituents.
A. glacialis sporangial (Type I) cells from the same and from different thalli germinate in situ to form annual germling populations, or to form a filamentous stage within the sporangium (Fig. 8E–H, arrows). Thalli with abundant sporangia (Type I) collected in a Ragged Channel embayment in late August early/September at 2–3 m, released zooids to form a large population of attached juvenile thalli (Figs 5A–H & 6D). Sporangium (Type I) contents develop thick walls with the greater number forming zooids after two mitotic (?) cell divisions, each cell liberating four zooids. A small number of the sporangia (Type I) cells maintain protoplasmic integrity and initiate erect, branching filaments (Fig. 8E, F & H, double white arrowheads). Branching of erect filaments begin early in their development with an anticlinal division of a terminal cell (Fig. 8G, white arrowhead). Paradoxically, cells of sporangial (Type I) filaments contain one belt-shaped, non-pyrenoidal plastid (Fig. 8H, arrows). Cells of germlings of all stages contain the multi, non-pyrenoidal plastids (Fig. 7C) a distinctive feature of A. glacialis. The role of these filaments in the species life history is unknown, possibly an over-wintering stage of A. glacialis, perhaps a source of vernal zooids. We posit that some of these immature thalli survive the following ice covered habitats to form a vernal population of the species. Growth of germlings formed from zooids released from Type I sporangia present on thalli in early September also developed numerous filamentous germlings and subsequent development of sporangial contents attached to presumably parental thalli. Elongate, non-expanded germlings (Fig. 7A) occurred in the sheltered, wave free environment at ca. 8–10 m. We believe that the difference in mature germling morphology at roughly comparable sizes (Fig. 6A vs. 7A) is possibly induced by the environment change where they occurred; wave swept in the former, mostly static water movement in the latter.
Kawai’s life history argument for synonymy and response
Synonymy is first based on nine shared genes with identical sequences (Kawai et al. 2015b). This followed by attempts to support the molecular data with previously published information concerning morphological and reproductive features of Platysiphon and Arcticophycus, and from observations of viable, but likely unavoidable stressed specimens Pl. verticillatus thalli under culture conditions. Response to the latter criteria as envisioned by Kawai et al. (2015b) follows. Our data on the growth and development of A. glacialis does not agree with the description cited by Kawai et al. (2015b) to change the Pl. verticillatus form to the A. glacialis form by the loss of the former species’ iconic Rat tail, viz. “… the distal thallus becomes lost to yield a foliose thallus form and stipe.” Suggested evidence of conspecificity cited by Kawai et al. (2015b) follows: “… parenchyma development throughout the entire Pl. verticillatus form.” We believe this is unlikely, as in situ observations of attached thalli retain a basal central tubular structure at the close of a current light season. There is no evidence of in situ tubular thalli becoming parenchymatous in late August/early September other than in the terete upper one-third of the thallus. Perhaps their comments are based on pre-senescent thalli in laboratory conditions.
The “… presumptive Platysiphon glacialis life history” cited by Kawai et al. (2015b, fig. 5) includes an endosporic gametophytic generation hypothesis (Henry 1984). Support for this hypothesis is presence of sporangial thick walled cells of presumed “gametophyte” ploidy. This critical premise does not fully agree with our own observations. Evidence to suggest a meiotic sequence is absent. In support of a sexual stage Kawai et al. (2015b, fig. 5g) claim “secondary zooids, presumptive gametes” fuse to form the “… juvenile stage of the terete thallus (Pl. verticillatus) with abundant hair-like assimilatory filaments.” Our evidence shows these thick-walled cells form asexual zooids or branched filaments in situ (see above). We have not observed cytological evidence of a sexual stage or that the two forms hybridize. Our field and laboratory evidence suggest an asexual life history accompanied by genetic recycling, similar to the suggested life history of Pl. verticillatus.
In support of this premise, the wide species occurrence of Pl. verticillatus from NW Greenland from one site in N Baffin Island, to the western Alaskan coast, and northern Labrador with no records of A. glacialis presence, and the distinctly different population development strongly suggests independent life histories. As a possible scenario that Arcticophycus is an asexual stage of Pl. verticillatus, our premise to the alternative is based laboratory cultures and empirical evidence. However, our reasoning includes ancestral lineage and evolutionary change from a distant form that incorporates features of the Halosiphon clade. More recent ancestors of both species were likely responsible for their current appearance after considerable time with accumulated environmental change. Gross change included separate attempts to adapt to an annual Arctic dark period. In addition to its distinctive vegetative features Platysiphon enigmatically evolved a cyst in its strategy of dark season survival. While on less solid evidence, we suggest Arcticophycus, while maintaining the robust upright stature of Halosiphon and Stschapovia, evolved a different over-wintering strategy to include two sporangial types. Change in both species, is attributed to evolutionary change from a common ancestor.
Significant questions remain. What role do the branched filaments of A. glacialis have in the life history of this species? Germlings formed from sporangium (Type I) are also pondered as a possible overwintering stage of the species life history. A third over-wintering structure can also be considered, the filamentous stage resulting from the germination of zooids from sporangium (Type II), the possible origin of vernal zoospores? Equally important is the future recognition of genetic unreported differences responsible for many distinctive, heritable, features that likely also separate Pl. verticillatus and A. glacialis.
Species with largely similar genomes differ when chunks of base pairs are changed by mutations that tweak their gene pools, gene shuffling, e.g., inverted in one species and not the other (Kirkpatrick 2010). Absolute certainty comes only through objective experiences. Our concepts of biology and those of phylogenetic analyses must inform the other. Despite genetic similarity, change of chromosome structure, gene on and off regulation and function may be largely responsible for some primate distinction viz. Pan spp. separation from Homo sapiens (Hu et al. 2012). Consider also the number of genes that are identical between chimpanzees and humans. Full genome comparisons, however, confirm (together with obvious morphological differences) that Pan troglodytes and Homo sapiens are different species. Taxon recognition is due to both shared and unshared genes or to diverse function of genes throughout the genome. Refined molecular protocol used to reveal these obvious differences between Pl. verticillatus and A. glacialis are yet to serve in support of the desynonyization of two arctic algal species. However, the lesson provided suggests that species difference may not be a function of additional genes but of rearranged genes, different expression of genes, how genes are expressed, or activated in diverse cell types. We suggest that over millennia, the late Tertiary/Pleistocene marine environments were the probable forcing cause for genetic inheritance in Pl. verticillatus and Pu. glacialis, and change from ancestral species. Reference to such changes are numerous, viz. Elias et al. (2012), Ostevik et al. (2012). A few to several genetic similarities (Zuccarello et al. 2017) and in a plethora of other studies, that cryptic “species” do not necessarily mean species synonymy. Science develops from a consensus of diverse concepts, those from a plurality of disciplines. Our efforts are those that tell the truth with the complexity of related facts. Can we really look askance, possibly ignore, the obvious evolutionary derived features that distinguish Pl. verticillatus and A. glacialis? It remains for coming questioning phycologists to determine whether these characters are sufficiently different at the molecular level when expressed in one taxon and not the other. For the present, we suggest that biological character expression in the field or laboratory is appropriate for species recognition.
Ample phylogenetic evidence (Kawai et al. 2015b) supports a Halosiphon Clade of four distinct, disparate species with a putative common ancestry (Pl. verticillatus, A. glacialis, H. tomentosus, and Stschapovia flagellaris). The oldest of the four taxa (Chorda tomentosa Lyngbye 1819 = Halosiphon tomentosus Jaasund 1957) provides genetic and taxonomic distinctiveness to the Halosiphon Clade. Further phylogenetic studies will likely identify the Clade ancestor as an erect brown alga in a developing north Polar Ocean prior to Pleistocene glaciation of mid to late Tertiary. The ancestor and its progeny must have evolved with the inclusion of the genetic and cytological components that characterize the Clade. But, when and how did the disparate distributions occur (viz. Stschapovia remains in the subarctic, NE Pacific Ocean, Platysiphon likely throughout the Arctic and NW Alaskan areas, Arcticophycus, likely throughout the Arctic and N Alaskan Pacific Ocean, the eastern Canadian Arctic and Greenland, while Halosiphon is ubiquitous throughout the Arctic, and boreal N Atlantic Ocean). Likely, it is only phylogenetic analyses that will further provide answers to these questions. Species of the Halosiphon Clade are appropriate as future research subjects owing to their unusual genetic similarities, their large biological differences, their current occurrences in cold Boreal to Arctic environments, and the posited common cool Boreal ancestor. It is also appropriate to suggest that three of the four species are remarkably amenable to unialgal culture. A. glacialis remains enigmatic as a tool in culture. Pl. verticillatus appears an especially ideal tool for studying phylogenetic forces that shape genomic characteristics and force species adaption to changing northern hemisphere marine environments. We support and identify Halosiphon Clade as a novel, concise group of four disparate, biological, and geographical wide spread, monogeneric taxa, collectively with promise to reveal answers that pertain to origins, relationships, even answers to species distribution patterns.
The flora of the Arctic Ocean is recognized (Wilce 2016), but origins of its species and those of the western N Atlantic Ocean remain questionable with several proposed paleocenographic origins (Lüning and Tom Dieck 1989, Wilce 1989). When, and on how many occasions, did species of the current Arctic flora adapt to alternating Boreal to Arctic temperature environments? What percentage of the western N Atlantic Ocean algal flora have their origin in the Polar Sea as opposed to a N Pacific Ocean (Maggs 1989, Van Oppen et al. 1995, Bringloe and Saunders 2017, 2018) or eastern N Atlantic Ocean origin (Lüning and Tom Dieck 1989, Wilce 1989). Species of the Halosphon Clade may assist in providing answers to these and related questions. In accordance with the above evidence, emended ordinal and family descriptions follow.
Stschapoviales H. Kawai in Kawai et al. 2015a; Wilce & Bradley, emended
Stschapovia flagellaris A. D. Zinova 1954, p. 242, figs 1–6 (3 sites sub-Arctic NW Pacific Ocean, northern Okhotsk Sea, Strait of Tartary). The order contains 3 families and 4 species: Stschapoviaceae (Kawai and Sasaki 2004), Stschapovia flagellaris (Zinova 1954), Platysiphonaceae (Kawai et al. 2015a), Platysiphon verticillatus (Wilce 1962), Arcticophycus glacialis (Rosenv.) Wilce & Bradley 2007, and Halosiphonaceae (Kawai and Sasaki 2001), Halosiphon tomentosus (Jaasund 1957).
Description
Erect thalli are simple, unbranched, slightly expanded, tubular, terete or linear thalloid; holdfasts are initially elongate saccate cells, later discoid or crustose; hairs are absent in erect thalli, photosynthetic assimilatory filaments in small clusters, often whorled, present in Platysiphon, Halosiphon, questionable in mature Stschapovia, lacking in A. glacialis; species in the order are conspicuously parenchymatous, excepting Pl. verticillatus with minimal parenchyma. Cortical cells contain non-pyrenoidal plastids and most significant, distinctive conservative genes are present in all species of the Stschapoviales.
Type species
Platysiphon verticillatus Wilce 1962.
Species description
Pl. verticillatus comparison with A. glacialis: mature thalli either less than 17 cm long, erect, partially tubular, weakly parenchymatous or larger (to 50 cm), erect, solid and entirely parenchymatous; delicate with numerous clusters of photosynthetic filaments or robust, naked, lacking photosynthetic filaments; growth in length from one or from several apical cells and from repeated cell divisions in the median thallus of each species; thalli slightly expanded in median portions with a terete, long, terminal “rat tail”-like distal region or blade uniformly flat, morphologically undifferentiated throughout; outermost cortical cells with numerous non-pyrenoidal plastids that vary in size and number within each species; reproduction with the formation of a chrysophyte-like cyst or with thick walled sporangia formed from cortical cells; species are sharply annual in occurrence with no cellular or ecological evidence of different species.
While Pl. verticillatus, H. tomentosus, and S. flagellaris are distinctive morphologically and reproductively. A. glacialis is distinctively unique in many respects but also, it can be characterized as the single recognizably undivided, conspicuous, non-kelp, brown algal thallus in the Arctic vegetation.
ACKNOWLEDGEMENTS
We are grateful for the critical reviews of J. R. Sears, J. F. Foertch, and J. H. Wilce, each contributed to manuscript development. We acknowledge the importance and instruction from papers concerned with assessment of species identity, such as Páll-Gergely (2017). That support was influential in strengthening our use of morphological criteria and in helping us to recognize that unlike genera, describing species without molecular support appears acceptable. We are especially grateful to D. J. Garbary for his scholarly persuasion in manuscript development and to M. J. Wynne for Latin usage. Monetary assistance is gratefully recognized. Support for several long periods of field work was provided by NSF (#0323015, P. I., R. T. W.), BIOS Project (Baffin Island Oil Spill), Department of the Environment, the Total Foundation, Paris, France, with F. Küpper in charge of the 2009 Ragged Channel field operations, for technical support from S. Brewer, Depart Biol, F. McIver, K. Kelly. Inform and Tech, Univ Mass, Amherst, Worcester State Univ, Worcester, MA, and B. I. Fuller (Graphic design).