Thermal and light impacts on the early growth stages of the kelp Saccharina angustissima (Laminariales, Phaeophyceae)
Article information
Abstract
Anthropogenic disturbances, including coastal habitat modification and climate change are threatening the stability of kelp beds, one of the most diverse and productive marine ecosystems. To test the effect of temperature and irradiance on the microscopic gametophyte and juvenile sporophyte stages of the rare kelp, Saccharina angustissima, from Casco Bay, Maine, USA, we carried out two sets of experiments using a temperature gradient table. The first set of experiments combined temperatures between 7–18°C with irradiance at 20, 40, and 80 μmol photons m−2 s−1. The second set combined temperatures of 3–13°C with irradiance of 10, 100, and 200 μmol photons m−2 s−1. Over two separate 4-week trials, in 2014 and again in 2015, we monitored gametogenesis, the early growth stages of the gametophytes, and early sporophyte development of this kelp. Gametophytes grew best at temperatures of 8–13°C at the lowest irradiance of 10-μmol photons m−2 s−1. Light had a significant effect on both male and female gametophyte growth only at the higher temperatures. Temperatures of 8–15°C and irradiance levels of 10–100 μmol photons m−2 s−1 were conditions for the highest sporophyte growth. Sporophyte and male gametophyte growth was reduced at both temperature extremes—the hottest and coldest temperatures tested. S. angustissima is a unique kelp species known only from a very narrow geographic region along the coast of Maine, USA. The coupling of global warming with high light intensity effects might pose stress on the early life-history stages of this kelp, although, as an intertidal species, it could also be better adapted to temperature and light extremes than its subtidal counterpart, Saccharina latissima.
INTRODUCTION
Large subtidal kelps are critical to near shore ecosystems. Kelps form habitat structure, nursery grounds, food for invertebrates and pelagic organisms, and buffer the shoreline from storms (Steneck et al. 2002, Smale et al. 2013). Macroalgal detritus fuels various food webs and is an important nutrient source for herbivores and detritivores (Krumhansl and Scheibling 2012, Duggins et al. 2016). In economic terms, these ecosystem services have been valued at billions of dollars annually (Kim et al. 2015, Rose et al. 2015, Krumhansl et al. 2016). Several kelp species are economically important for food and alginates (Kim et al. 2017, Wells et al. 2017) including Saccharina japonica in Asian countries. In the North Atlantic of the USA, the key species used for the developing seaweed aquaculture industry is Saccharina latissima, or sugar kelp.
Sea-surface temperature (SST) is a primary factor driving macroalgal biogeographic distributions in the marine intertidal and, along with light and nutrient requirements, confine macroalgae to the shallow, photic zones (Lüning 1990, Adey and Steneck 2001, Steneck et al. 2002). Evidence is growing that kelp habitats are under threat from multiple stressors including warming SST temperatures, pollution, overharvesting and other anthropogenic disturbances that combined can have major consequences for the structure and function of near-shore coastal ecosystems (Smale et al. 2013, Krumhansl et al. 2016). Under some current climate change scenarios, kelps appear to be at risk in some environments; therefore, it is critical to recognize the photosynthetic responses of kelp to temperature variations to better understand resilience of these important habitat-forming species (Colvard et al. 2014, Krumhansl et al. 2016). In the past few decades, studies have shown the retreat of northern hemisphere kelp beds northward with small fluctuations in SST (Fernández 2011, Pereira et al. 2011, Moy and Christie 2012, Wernberg et al. 2016). Even more studies show kelp declines due to warming (Díez et al. 2012, Harley et al. 2012, Bartsch et al. 2013, Voerman et al. 2013, Krumhansl et al. 2016, Park et al. 2017). The Gulf of Maine (GOM) in the northwestern Atlantic showed unprecedented warming in the 2012 ocean heat wave with temperature anomalies of ~2°C above the annual 1998–2011 climatology (Mills et al. 2013). Furthermore, GOM data shows SST increasing faster than 99% of the global ocean over the past decade (Pershing et al. 2015). This warming has critical implications for near shore flora and fauna (Lesser 2016), with a northwest migration already observed for some local finfish species (Mills et al. 2013). Furthermore, future climate scenarios, predict global temperature increases of ~2°C by 2100, assuming significant climate action and low-emission scenarios (RCP3-PD), while high-emission, non-mitigating future scenarios (RCP8.5) predict increases of up to 6.1°C (Rogelj et al. 2012). This is especially critical for intertidal organisms exposed to desiccation stresses with the ebbing tides.
Kelps have a two-stage alternation of heteromorphic generations with a prominent sporophytic blade phase giving rise via meiosis to a microscopic filamentous gametophytic phase. These microscopic stages are critical for the maintenance of the population after annual sporophytes decay (Hoffmann and Santelices 1991, Pereira et al. 2011). It has been shown that the gametophytic stages of S. latissima populations from the western North Atlantic have an upper critical thermal limit of 22°C, while gametogenesis was suppressed at temperatures above 17°C (Redmond 2013). Similarly, S. latissima juvenile sporophyte development was inhibited at 17°C and the upper temperature limit for growth was between 18–19°C (Kain 1969).
The light saturation point of growth for S. latissima juvenile sporophyte blades has been shown to be at 50–70 μmol photons m−2 s−1, with an optimum photon flux density of 110 μmol photons m−2 s−1 as the sporophytes approach 6 weeks old (Kain 1969, Egan et al. 1989). Reports for the same species indicate that 1- to 2-year-old fronds show photoinhibition at 250 μmol photons m−2 s−1 (Fortes and Lüning 1980). Wiencke and Fischer (1990) showed that Laminaria digitata adult sporophyte growth was light saturated between 55 and 105 μmol photons m−2 s−1. S. latissima gametophytes grow best at low light levels of 5–20 μmol photons m−2 s−1 (Lee and Brinkhuis 1988) up to 25–50 μmol photons m−2 s−1 (Egan et al. 1989).
A close relative of S. latissima, Saccharina angustissima (Collins) Augyte, Yarish & Neefus is an ecologically, morphologically and genetically distinct kelp found in the GOM (Augyte et al. 2018). Its range spans only 8 nautical miles in the low intertidal on ledges and islands exposed to high wave forces (Mathieson et al. 2008, Mathieson and Dawes 2017, Augyte et al. 2018). The observed recorded SST increases in the GOM and future climate change predictions coupled with the limited distributional range of this rare kelp, place it in danger of extinction from its current habitat. Therefore, the aim of this study was to identify ideal temperature and light conditions for the early developmental stages of this economically important kelp species, S. angustissima (Collins) Augyte, Yarish & Neefus from Casco Bay, Maine, USA.
MATERIALS AND METHODS
Two independent experiments tested the effects of temperature and light on the early life stages of S. angustissima, specifically gametogenesis, gametophyte growth, and early sporophyte development. The first set of experiments (in 2014) used five temperatures between 7 ± 1 and 17 ± 1°C (at 7, 9, 12, 15, and 17°C), and irradiance of 20, 40, and 80 μmol photons m−2 s−1. The second set of experiments (in 2015) had five temperatures between 3 ± 1 to 13 ± 1°C (at 3, 5, 8, 10, and 13°C), and 3 irradiance levels set at 10, 100, and 200 μmol photons m−2 s−1. The photoperiod was set to 12 : 12 light : dark cycle in both experiments.
Sorus tissue was collected from mature blades located in the low intertidal from Bailey’s Island, Harpswell, ME (43°43.32′ N, 69°59.46′ W) during the extreme low tides of October 2014, and again in October 2015. Fall is the peak reproduction period of S. angustissima when about 85–95% of the blades are covered with sorus tissue (Philibert 1990). The sorus tissue was cut, cleaned with an iodine wash, wrapped in paper-towels and allowed to desiccate overnight. The following day the tissue released meiospores when placed in sterile seawater. The resulting concentration of the meiospore solution was, on average, 135,000 meiospores mL−1. Two drops of this meiospore solution (~27,500 meiospores) were pipetted onto glass cover slips and allowed to germinate in a moisture chamber at 10°C in complete darkness, following Egan et al. (1989). After settlement of 48 h, the cover slips were placed into 300 mL crystalline Pyrex dishes with 250 mL of sterilized seawater enriched with 0.5× Provasoli’s Enriched Seawater (Provasoli 1968) medium, which was replaced weekly. Each light level and temperature treatment (n = 15) had 3 pseudo-replicates for a total of 45 dishes on the gradient table. Temperature measurements were taken with a digital thermometer (Traceable; Control Co., Webster, TX, USA) and light was measured with a Li-Cor LI-1000 (Li-Cor, Inc., Lincoln, NE, USA) photometer. Each week, for a total of 4 successive weeks, a cover slip was randomly chosen from each light/temperature treatment dish and photographed with a camera (PixeLINK, Rochester, NY, USA) attached to a compound microscope. Since 100 gametophytes or sporophytes were counted from each treatment, every coverslip was considered a replicate. Cover slips were discarded after use. Images of meiospore germination, gametophyte and sporophyte length and size were analyzed using ImageJ software (Abràmoff et al. 2004).
Germination was recorded during week 1 as presence/absence data with the observation of germ tube formation, indicating that meiospores had settled and were developing into gametophytes. Sex of developing gametophytes was distinguished based on size and diameter of cells—females were clearly bigger with plumper cells while male filaments were smaller with thinner cells. The female gametophytes were measured by counting the number of cells in each filamentous multicellular structure in week 3 of the experimental treatments. The male gametophytes were measured by taking the length of the longest part of each filamentous multicellular structure at week 3 of all the experimental treatments. Length was used instead of number of cells because the individual male cells were small and indistinguishable within the multicellular filaments. Finally, total length of juvenile sporophytes was measured at week 3 and again at week 4 of each experiment.
Separate data analyses were carried out for the two independent experiments. For count data of kelp spore germination, a chi-square test of independence was run to test if germination at different light levels was dependent on temperature and vice versa.
The effects of light and temperature on the measured responses (size and length) were analyzed by running separate two-way ANOVAs using the generalized linear model with R software (R Core Team 2013). The Levene’s test was run to check equal variance and the Shapiro-Wilk test for normality or residuals. All assumptions for normality and equal variance were met. Significance was defined as p-value < 0.05. When testing statistical differences between irradiance and temperatures, averages were taken for gametophytes and sporophytes that were counted on one coverslip and each coverslip was treated as a replicate. Afterword, significant differences between treatments were found a posteriori with Student-Newman-Keuls (SNK) test using the R package “agricolae.”
RESULTS
Germination rates
For the higher temperature experiments with ranges 7–17°C (from 2014), germination rates were over 90% for all treatments except for the temperatures 12 and 15°C exposed to the highest light levels tested of 80 μmol photons m−2 s−1 (Fig. 1A). During both years of experiments, a significant interaction was found between light and temperature on germination rates (2014; chi-squared = 59.82, p < 0.001, 2015; chi-squared = 131.44, p < 0.001) (Fig. 1A & B).
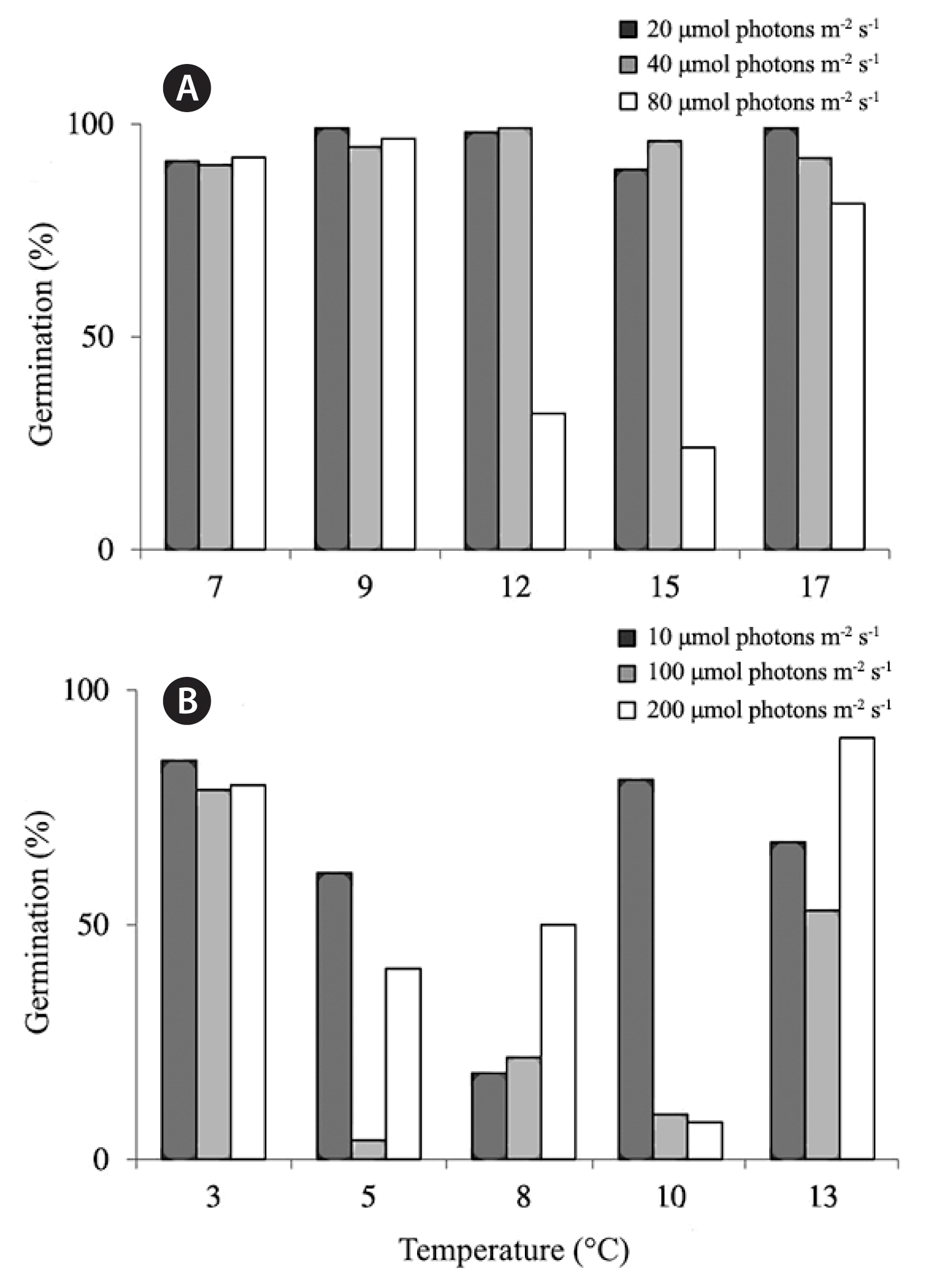
Results for the 2014 (A) and the 2015 (B) experiments of kelp germination rates after settlement exposed to different temperature and light treatments. Legend indicates light level as μmol photons m−2 s−1.
Female gametophyte size
In the higher temperature experiments (from 2014), temperature (p = 0.0259) (Table 1, Fig. 2A) but not light (p = 0.3572) were found to significantly affect female gametophyte size. Overall, except for the lowest lows and the highest highs, the sizes of the female gametophytes were very similar and indistinguishable between treatments. The highest mean cell numbers were found in treatment 17°C at 40 μmol photons m−2 s−1 with 3.3 (±0.6) cells (Fig. 2A). The other treatments ranged in mean cell numbers of 1.2–2.7.

Two-way ANOVA table showing statistical significant relationships between Saccharina angustissima gametophyte or sporophyte size with light and temperature as explanatory variables
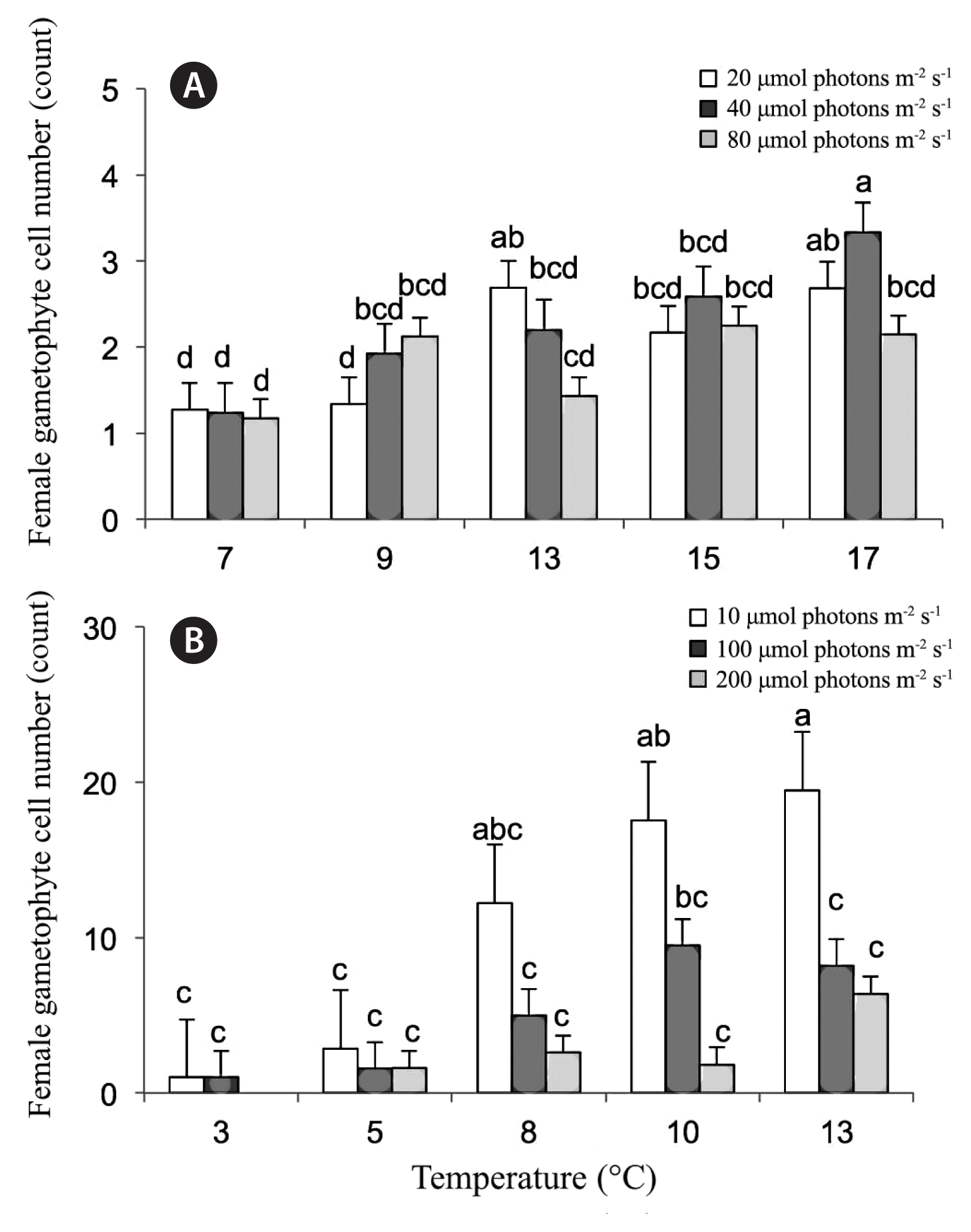
Results for the 2014 (A) and the 2015 (B) experiment on the effects of temperature and irradiance on cell number of female gametophytes in week 3 (means ± standard error). Legend indicates photon fluence rates (μmol photons m−2 s−1). Treatment combinations with no letters (a–e) in common were found to be significantly different by Student-Newman-Keuls analysis.
In the second set of experiments with lower temperatures ranges of 3–13°C (from 2015), both light (p = 0.0194) (Table 1, Fig. 2B) and temperature (p = 0.0253) were found to have significant effects on female gametophyte size. The largest female gametophytes were found at the lowest light levels of 10 μmol photons m−2 s−1 with mean number of cells averaging at 12.2 ± 4.3, 17.6 ± 3.2, and 19.5 ± 5.3 for temperatures 8, 10, and 13°C, respectively. The coldest temperatures (3 and 5°C) showed low growth at all light levels. Growth was not observed at the coldest treatment (3°C) coupled with the highest light level (200 μmol photons m−2 s−1). While year to year variation was not tested in the analysis, it was observed that overall, female gametophytes grew larger than in the first set of experiments.
Male gametophyte size
For the higher temperature experiments (from 2014), temperature had a slight significant effect on male gametophyte length (p = 0.052) (Table 1, Fig. 3A) while light was not significant (p = 0.770). The largest male gametophytes were found at 15°C at 40 and 80 photons m−2 s−1 with lengths of 111.6 ± 13.9 and 91.0 ± 17.0 μm, respectively and at 17°C at 20 photons m−2 s−1 with a length of 83.4 ± 12.7 μm. All other means of the other treatments were not significantly different from each other with average ranges of 12.6–54.0 μm.

Results for the 2014 (A) and the 2015 (B) experiment on the effects of temperature and irradiance on filament length (μm, means ± standard error) of male gametophytes in week 3. Legend indicates photon fluence rates (μmol photons m−2 s−1). Treatment combinations with no letters (a–e) in common were found to be significantly different by Student-Newman-Keuls analysis.
In the second set of experiments (from 2015), neither temperature (p = 0.344) (Table 1, Fig. 3B) nor light (p = 0.213) were found to have a significant effect on male gametophyte length. At 8°C, a spike in the length of gametophytes grown at 10 μmol photons m−2 s−1 of 135 ± 13.4 μm was observed. The second largest size sets were again measured at the lowest light level tested at temperatures of 10 and 13°C with lengths of 75.6 ± 6.6 and 85.6 ± 8.4 μm, respectively. The rest of the means were not significantly different from each other.
Sporophyte growth
For the first set of experiments (from 2014), some sporophyte development was already observed during week 2, but measurements only started in week 3. Both light and temperature were found to have significant effects of sporophyte length only in week 4 not in week 3 (week 4: light, p = 0.032; temperature, p < 0.001; week 3: light, p = 0.8966; temperature, p = 0.0977) (Table 1, Fig. 4A & B). In both weeks 3 and 4, the largest sporophytes were found in treatment of 12°C and 40 μmol photons m−2 s−1 at mean lengths of 186 ± 14 μm in week 3, growing to lengths of 557 ± 39 μm by week 4. In week 4, large sporophytes were also measured at 15°C and 40 μmol photons m−2 s−1 conditions with lengths of 536 ± 40 μm. At 17°C, only a few sporophytes were produced under the lowest light levels, while the other light conditions produced only gametophytes. The coldest temperature tested, 7°C, produced very small sporophytes.
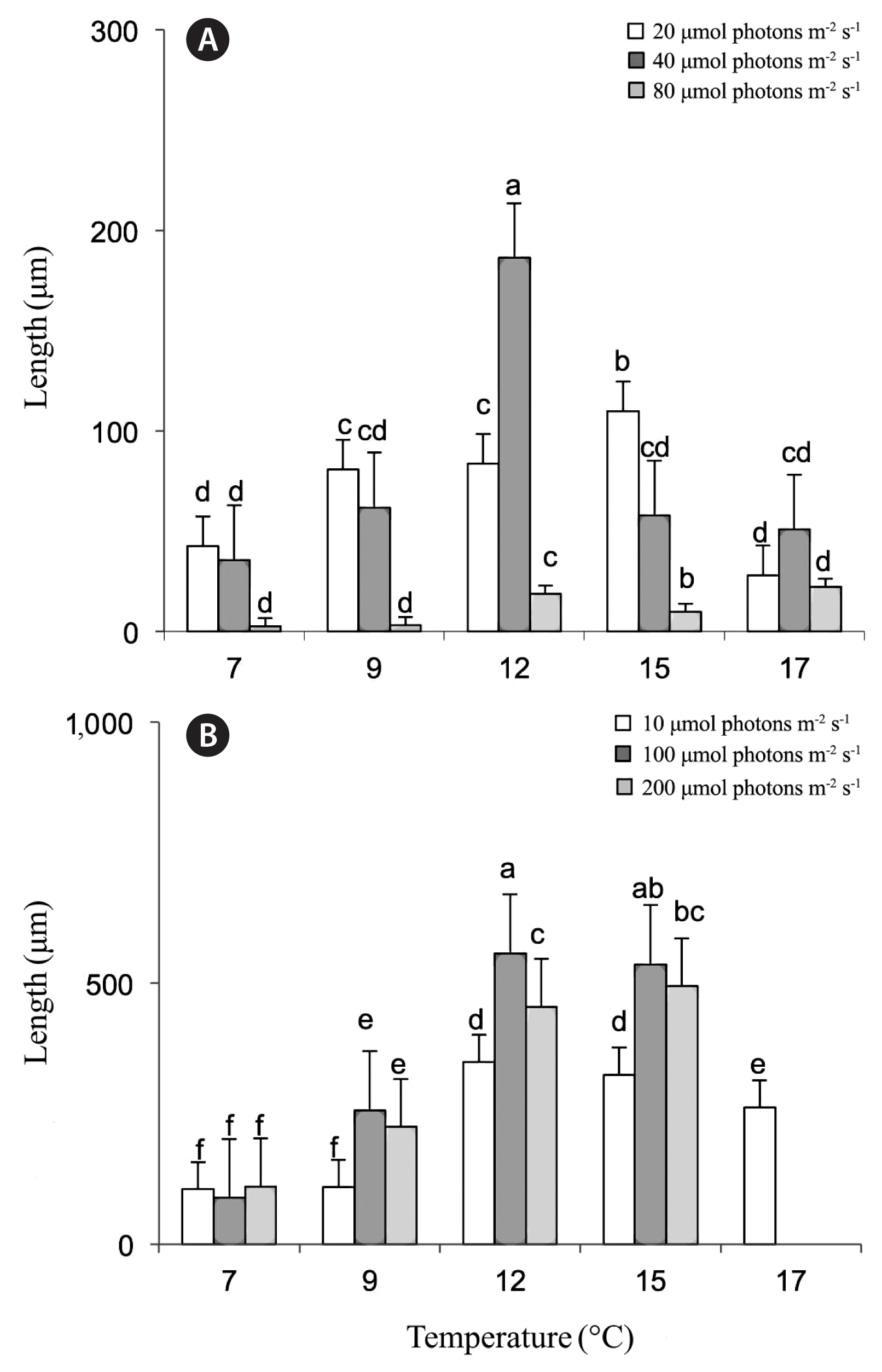
Effects of temperature and irradiance on kelp sporophyte lengths (μm, means ± standard error) during 2014 in treatment 7–17°C for week 3 (A) and week 4 (B). Legend indicates photon fluence rates (μmol photons m−2 s−1). Treatment combinations with no letters (a–e) in common were found to be significantly different by Student-Newman-Keuls analysis.
For the second set of experiments (from 2015) light but not temperature were found to exert a significant effect on sporophyte length in both weeks 3 and 4 (week 3: light, p = 0.0398; temperature, p = 0.8508; week 4: light, p = 0.0373; temperature, p = 0.2385) (Table 1, Fig. 5A & B). In week 3, only gametophytes, no sporophytes, were observed at the following conditions; 3°C for 200 μmol photons m−2 s−1 and 10 and 13°C for 10 and 200 μmol photons m−2 s−1. Longest sporophytes were found at 10°C at 100 μmol photons m−2 s−1 at mean lengths of 197 ± 19 μm. In week 4, there were no sporophytes, only gametophytes, observed for the highest light treatments tested of 200 μmol photons m−2 s−1. Treatments of 8 and 13°C at 100 μmol photons m−2 s−1 produced the largest sporophytes at 523 ± 24 and 650 ± 18 μm, respectively. Data is missing for week 4, temperature treatment 10°C at 100 μmol photons m−2 s−1 so it is unclear if the results from week 3 would have carried over.
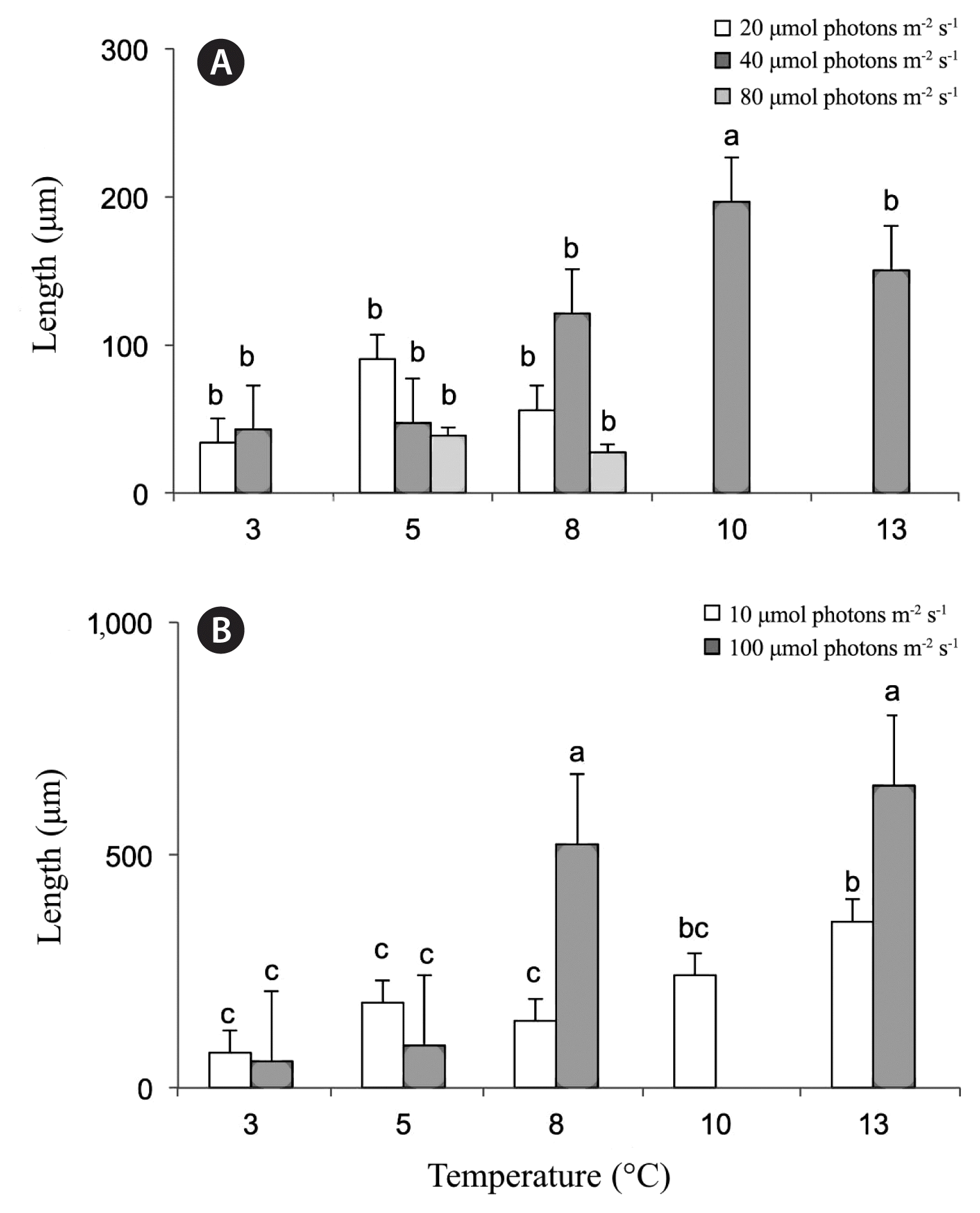
Effects of temperature and irradiance on kelp sporophyte lengths (μm, means ± standard error) during 2015 in treatment 3–13°C for weeks 3 (A) and 4 (B). Legend indicates photon fluence rates (μmol photons m−2 s−1). In week 3, there were no observable sporophytes, only gametophytes in 10 and 13°C at 10 and 200 μmol photons m−2 s−1. In week 4, there were no observable sporophytes in light treatment 200 μmol photons m−2 s−1 with all temperature treatments tested. Data is not available for week 4, temperature treatment 10°C at 100 μmol photons m−2 s−1. Treatment combinations with no letters (a–e) in common were found to be significantly different by Student-Newman-Keuls analysis.
DISCUSSION
Kelp meiospores and gametophytes are particularly vulnerable to fluctuations in environmental conditions (Müller et al. 2008, Zhang et al. 2013) and it is therefore critical for kelp physiology and biogeography to understand how these microscopic phases respond in light of changing climate scenarios. In general, high germination rates (90%) of kelp of meiospores were observed at temperatures ranging from 7–17°C in light conditions of 20–80 μmol photons m−2 s−1 except for at 12 and 15°C. In the second experiment, the pattern for germination rates is not as obvious and warrants further study.
In the low intertidal in Casco Bay, Maine, Saccharina angustissima reaches its peak in sorus and thus meiospore production and release in October to November (Philibert 1990, personal observation) when SST are at 10–13°C (Fig. 6A). The coldest air temperatures follow during January through April dropping to lows of 4 ± 2°C (Fig. 6B). In this study, the coldest temperatures tested at 3–5°C, showed trends of a reduction in size of male and female gametophytes, regardless of light level. One-celled female gametophytes still produced an egg that allowed fertilization to occur as sporophytes were observed in the subsequent week in those conditions. Furthermore, a colder tolerance by female gametophytes was observed, which in the field, may translate to earlier growth rates for females compared to males. Previous work shows that S. latissima female gametophytes may be more sensitive to increases in temperature, and higher temperatures may produce more male than female gametophytes (Lee and Brinkhuis 1988, Egan et al. 1989). For the kelp Laminaria ochroleuca, low light levels and temperatures of 10–12°C were ideal for producing female gametophytes of more than 20 cells in the warm temperate species (Price et al. 2003). While gametophyte development of the cold temperate kelp, Saccorhiza polyschides, in Portugal was not observed at very high temperatures of 25°C (Pereira et al. 2011).
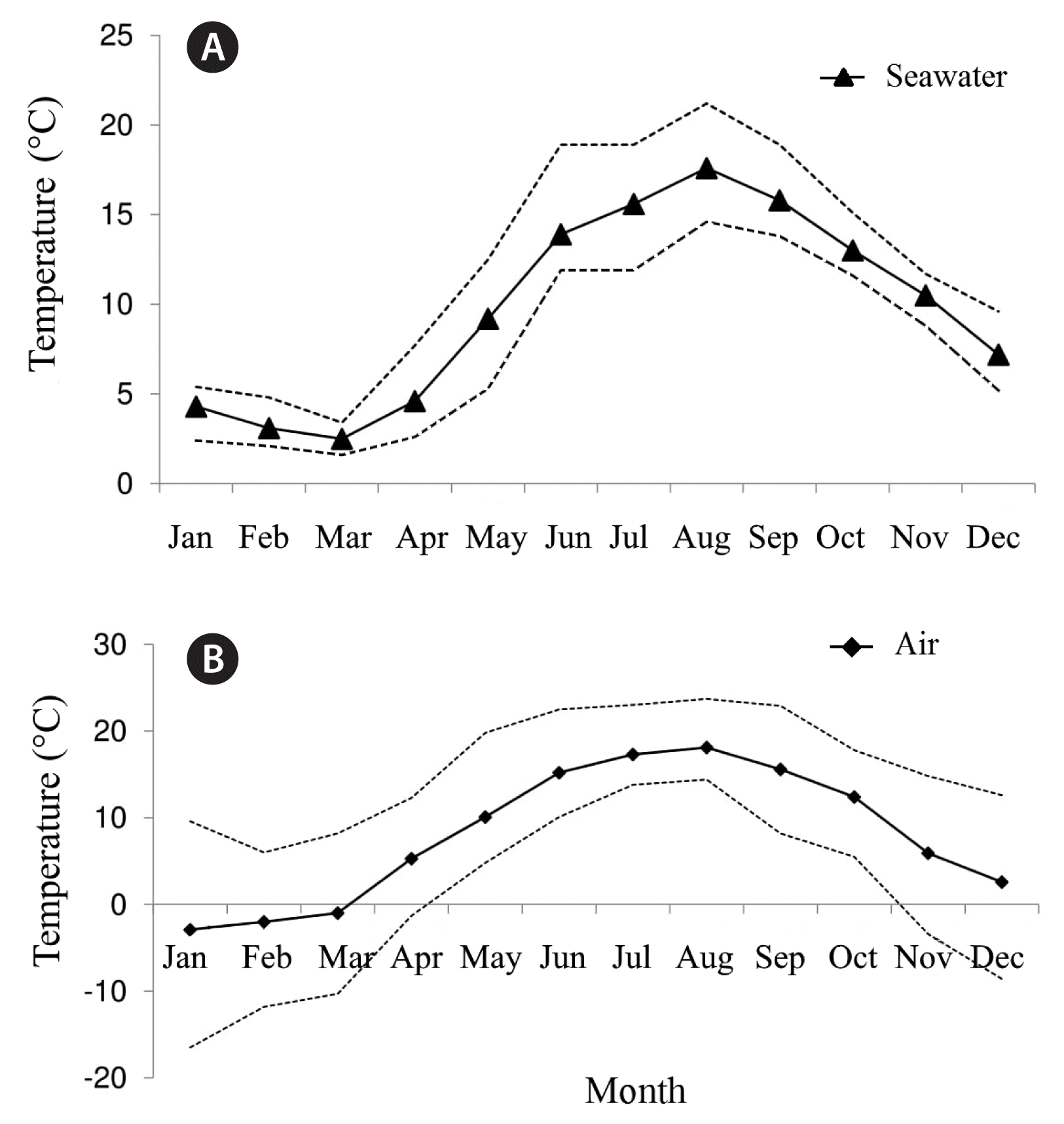
Monthly seawater (A) and air (B) temperature (°C) averages for 2014 for Casco Bay, Maine. Dashed lines represent average high and lows for the months. Data obtained from NERACOOS buoy 44007.
In the field, the S. angustissima kelp gametophytes must have temperature triggers that stimulate growth when SST reach above 7°C. This has implications for future reproductive success of this species in relation to climate change. For example, if SST warming of a few degrees occurs during the winter, the gametophytes might have a competitive advantage in production of biomass, especially under low light conditions. Yarish et al. (1990) reported a reduction in gametangia and sporophyte production and development of Saccharina longicruris at 5°C, especially under high photon fluence rates (>40 μmol photons m−2 s−1), but inter-annual variation has also been reported (Egan et al. 1989). In this study, longer female gametophyte filaments of 18–19 cells were measured in some of the warmer conditions at 10 and 13°C; temperatures that would only be observed in the field starting in May. Mature S. angustissima sporophytes have been collected at the site in June, but it is unknown how early in the season blade development commences. According to typical kelp life cycle phenology, spores settle within hours to a few days of release, gametes are produced over the next week and fertilization occurs shortly after leading to the development of a zygote and eventually to a macroscopic sporophyte (Reed et al. 1992, Schiel and Foster 2006). For S. angustissima growing in the high wave environment, it is predicted that meiospore settlement occurs rapidly after release and that fertilization of gametes soon after. However, additional research is needed to identify the mechanisms for meiospore settlement and fertilization success including for how long, if at all, filamentous gametophyte banks persist over the winter into spring. Our results confirm that optimal S. angustissima gametophyte growth in culture conditions is at temperatures ranging 8–13°C with light levels from 10 to 100 μmol photons m−2 s−1 suggesting that if gametophytes persist over the winter, they are able to grow into large filaments when both air and water temperatures are closer to their optima observed in lab experiments.
Kelp gametophytes have the capacity to withstand long periods of darkness and thus have exceptionally low-light demands (Bartsch et al. 2008). Laminaria digitata, L. ochroleuca, and Laminaria hyperborea gametophytes become fertile at irradiance as low as 2–4.5 μmol photons m−2 s−1 (Tom Dieck 1992). In this study, the lowest light level tested, 10 μmol photons m−2 s−1, produced some of the highest growth for gametophytes. Female gametophytes showed some growth and development at the highest light levels, 200 μmol photons m−2 s−1, in all but at the coldest (3°C) condition. However, the highest light condition was detrimental to some of the male gametophytes and these grew only under the temperature conditions of 8 and 13°C. Male gametophytes, therefore, are able to tolerate some high light but only when coupled with their optimal temperature conditions. Previous studies document light saturation at 70 μmol photons m−2 s−1 for S. latissima (Fortes and Lüning 1980). Similarly, in this study, high light was detrimental to juvenile S. angustissima sporophyte development. While in week 3 some sporophytes were observed at the highest light tested (200 μmol photons m−2 s−1) at 5 and 8°C, by week 4 there were no sporophytes observed under these light conditions. However, light levels of 100 μmol photons m−2 s−1 still produced large sporophytes. The intertidal kelp, S. angustissima might be better adapted to higher light levels than the subtidal S. latissima and thus might have a competitive advantage succeeding in future scenarios when facing changing environmental conditions.
The largest sporophytes were observed at temperatures 12 and 15°C for the first set of experiments and again at 13°C for the second set. The highest temperature tested of 17°C showed slight sporophyte development in week 3 for all light levels, but by week 4 only at the lowest light level. Therefore, high temperatures coupled with high light levels were detrimental to juvenile sporophyte growth and development. Within its current restricted distribution, in Casco Bay, Maine, sea-surface and air temperatures above 17°C are experienced by fully developed adult kelp sporophytes in late summer for brief periods during emersion. During the months of August and September, the SST averages are up to 17 ± 3°C, while the air temperature reaches highs of 18 ± 6°C (Fig. 6A & B). For both experiments, the cold temperature treatments (3–7°C) produced stunted sporophytes. Literature shows a broad range of optimal temperatures for S. latissima sporophyte growth of 10–15°C in temperate regions (Fortes and Lüning 1980, Yarish et al. 1990) to as low as 5–10°C growth optima in Arctic regions (Park et al. 2017). In Alaska, decreased settlement and growth was observed for S. latissima spores and gametophytes when exposed to temperatures of 21°C and low salinity, however, the kelp was overall resilient to temperature changes even when these were extreme and abrupt (Lind and Konar 2017). In the field, an additional stress for S. angustissima is exposure to below freezing air temperatures during winter low tides that may drop to as low as −2°C (Fig. 6B). The results presented in this study show that sporophyte development was stunted at the coldest and hottest temperatures tested alluding to an optimal temperature for growth. The optimal temperature range was found to be 8–15°C. Specifically, temperature and light conditions for sporophyte development were 12 and 15°C at both 40 and 80 μmol photons m−2 s−1 and at 8 and 13°C at 80 μmol photons m−2 s−1.
It is critical for kelp to be able to acclimate to short-term heat stress and seasonal temperature shifts, especially for kelp near their physiological temperature limits (Wernberg et al. 2010, Bartsch et al. 2013). There is little experimental evidence suggesting that increasing SSTs will allow species to evolve fast enough to stay within current geographic zones because of global climatic change (Parmesan 2006). The kelp S. angustissima appears to have limited dispersal capability and grows in a radius of eight nautical miles (Mathieson et al. 2008, Mathieson and Dawes 2017, Augyte et al. 2018). This study indicates that temperatures above 15°C inhibit S. angustissima sporophyte formation, especially at high irradiance and thus, warming SST could limit its ability to retreat northward if it becomes pushed out of its optimal temperature range in the Gulf of Maine if the current warming trend of (0.23°C y−1) continues as has been observed in the last decade (Pershing et al. 2015). Overall, the filamentous gametophytes were shown to be more tolerant of higher temperatures than juvenile sporophytes especially when coupled with high irradiance. Warming temperatures could thus shift reproduction times to earlier in the season and as well as lead to more observed production during the spring and fall seasons as opposed to the summer months when this kelp is at its current peak blade production.
ACKNOWLEDGEMENTS
A special thanks to N. Ramakrishna, A. Ferro, and Y. Huo for assistance with gradient plate experiments, A. Lamb and J. K. Kim for the review of manuscript, and J. Kübler for statistical expertise. This work was supported by The Maine Aquaculture Innovation Center (Project No. 15-11), The Francis Rice Trainer Endowment Fund to the Dept. of Ecology and Evolutionary Biology and the Connecticut Museum of Natural History, Sigma Xi Grants-in-Aid of Research, The Woman’s Seamen’s Friend Society of Connecticut, and USDA/National Institute of Food and Agriculture (Project Award # 2014-70007-22546).