INTRODUCTION
Over the past five years, an unrecognized diversity of aquatic oomycetes has been unveiled by many independent studies using field observations, metabarcoding, single-cell isolation of phytoplankton, and even time-consuming laboratory cultures. To date, these organisms appear to form a hugely diverse group of intracellular holocarpic parasites that infect marine and freshwater algae: green algae (Chlorophyta), red algae (Rhodophyta), brown algae (Phaeophyceae), and diatoms (Bacillariophyceae). They have been suggested to play potentially important roles in controlling phytoplankton dynamics (Garvetto et al. 2018) and represent a known or potential threat for commercially cultivated algae (Badis et al. 2019, 2020). The severe economic costs of these pathogens have increased with the spread and intensification of seaweed (esp. red algal) aquaculture over the last decades (Kim et al. 2014, Wen et al. 2023).
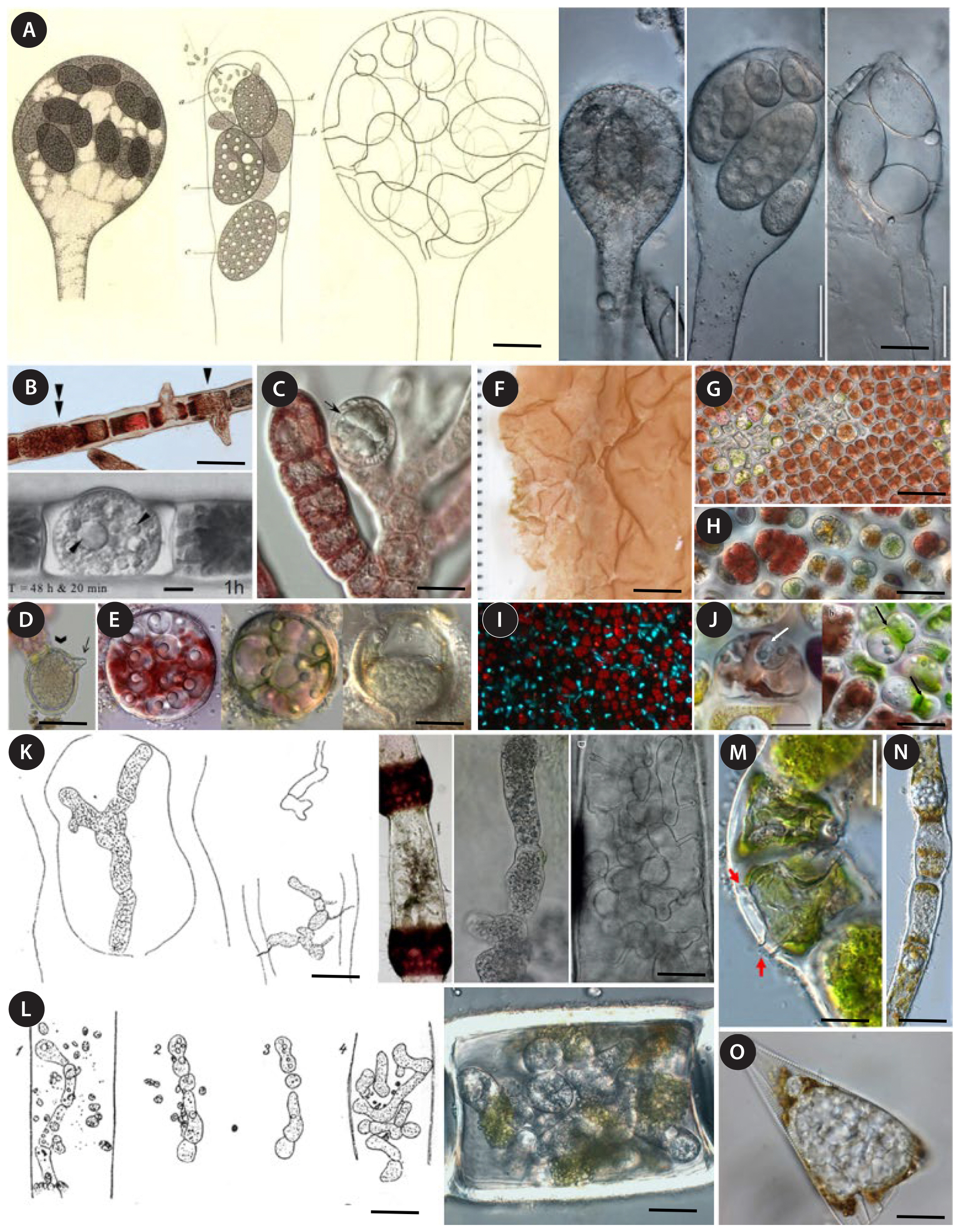
These intracellular, obligately parasitic oomycetes infecting algae have started being described in the second half of the 19th century (Fig. 1), with the delineation of taxa based on a combination of morphological characters (e.g., flagellation of zoospores) and life cycle (e.g., morphogenesis of sporangia). These early studies also suggested that host specificity could have taxonomic utility, yet this needed further study (Sparrow 1960). Later studies on laboratory cultures indicated that parasites had greater host diversity than originally assumed from field observations (Sekimoto et al. 2008, 2009, Fletcher et al. 2015, Klochkova et al. 2016, 2017). Until the first molecular sequences became available at the turn of this century, the phylogenetic position of these oomycetes has remained all but uncertain, and some have long been misidentified as chytrids or even hyphochytrids (Gachon et al. 2017).
Olpidiopsis was originally described as an endobiotic parasite of various oomycetes and freshwater and marine algae (Cornu 1872, Sparrow 1960). The type species is O. saprolegniae (A. Braun) Cornu, a parasite of Saprolegnia spp. Recently, a parasite infecting Saprolegnia parasitica was isolated and barcoded, this host species matched the original host (Saprolegnia spp.) (Braun 1855) and its pathogen matched O. saprolegniae (Cornu 1872), leading to its designation as an epitype (Buaya et al. 2019). Several holocarpic pathogens of red algae have historically also been placed in the genus Olpidiopsis, due to their morphological similarity with holocarpic pathogens of oomycetes. They share a usually single spherical holocarpic zoosporangium, with one or several evanescent discharge tubes (Sparrow 1960), yet appear to be phylogenetically distant from the epitype of O. saprolegniae (Buaya et al. 2019). These species of red algal parasites for which 18S rRNA data is available, widely known as Olpidiopsis bostrychiae, O. feldmanni, O. heterosiphoniae, O. muelleri, O. palmariae, O. porphyrae, and O. pyropiae, therefore cannot be included in this genus any more (Buaya et al. 2019, 2023a, Buaya and Thines 2020).
Another intracellular pathogen of red algae is Pontisma lagenidioides H. E. Petersen (1905), which forms two types of sporangia: simple “olpidiod” sporangia, and sporangia stemming from fragmenting thalli (Petersen 1905, Sparrow 1960). Recently, a parasite of an alga identified as Ceramium rubrum which had a “thallus […] usually composed of a series of somewhat irregularly cylindrical, sausage-like segments” was sequenced and used to epitypify the genus Pontisma (as P. lagenidiodes). Following an earlier speculation by Dick (2001), these authors then transferred all holocarpic parasites of red algae into the genus Pontisma though this clade was unsupported in their phylogenetic analyses (Buaya et al. 2019, Buaya and Thines 2020).
Sirolpidium H.E. Petersen was originally erected alongside Pontisma to account for the ability of its type, S. bryopsidis to differentiate two types of thalli (olpidioid and fragmenting) within its green algal host Bryopsis plumosa (Petersen 1905, Sparrow 1960). The morphogenetic similarities between Pontisma and Sirolpidium had long been recognized and led to the suggestion of merging both genera (Karling 1942). Recently, a pathogen of the green filamentous Ulotrichales, Capsosiphon fulvescens, tentatively likened to Sirolpidium bryopsidis, despite its apparent inability to infect Bryopsis plumosa and different geographic origin, was found to have a partial 18S sequence identical with Pontisma lagenidioides (Buaya et al. 2021). Finally, a novel pathogen of a second green filamentous Ulotrichales, Urospora neglecta, which only showed olpidioid thalli and had a 18S sequence that was sister, though with only moderate bootstrap support, to the red algal parasite Olpidiopsis porphyrae. Having realized that Karling retained Sirolpidium over Pontisma, these authors now propose to assign all holocarpic, pathogens of red and green algae mentioned above to the genus Sirolpidium, within the monogeneric order Pontismatales (Buaya et al. 2023b). As defined in the latter paper, this group also encompasses pathogens of diatoms and brown algae (Garvetto et al. 2018, Buaya and Thines 2022), as well as environmental sequences from uncharacterized organisms.
This body of work has led to repeated name changes to some important red algal pathogens twice over the past four years (Buaya et al. 2019, 2023b), though it was obvious from all these papers that this new circumscription of Pontisma (or lately, Sirolpidium) and the order Pontismatales was unsupported in any analyses (e.g., Badis et al. 2019, Buaya et al. 2019, 2021, 2023b, Buaya and Thines 2020). We decided to reinvestigate the phylogeny of this group based on the same data set of publicly available small subunit ribosomal DNA (SSU rDNA). Our results confirm that the taxa Pontisma, Sirolpidium, and Pontismatales are not supported. We discuss the negative consequences of premature nomenclatural changes to stakeholders and make a recommendation to conserve incorrect, yet widely understood, original names of red algal pathogens until a supported phylogeny can be obtained and adequate nomenclatural changes made.
MATERIALS AND METHODS
The nuclear encoded small subunit ribosomal 18S RNA gene (SSU) dataset was copied Buaya and Thines (2020) with added all recent sequences of holocarpic biotrophic algal parasites (e.g., Buaya et al. 2023b) from GenBank. Several alignment methods were used, that all employ different algorithms and penalty scores: (1) Pagan using the Was@bi webserver (http://was.bi/) and default parameters (Löytynoja 2021); (2) PRANK alignment using the stand alone package also incorporating the default parameters (Löytynoja and Goldman 2010); (3) T-Coffee for multiple DNA sequence alignment (M-Coffee) (Notredame et al. 2000) used the webserver (https://tcoffee.crg.eu/apps/tcoffee/) (Di Tommaso et al. 2011); (4) the SINA aligner (Preusse et al. 2012) on the SILVA website (https://www.arb-silva.de) (Quast et al. 2012); and finally, (5) we used MAFFT with the alignment strategy Q-INS-I, on the online server (https://mafft.cbrc.jp/alignment/server/) (Katoh and Standley 2013, Katoh et al. 2019).
Maximum-likelihood analyses were implemented using IQ-TREE 2 (Minh et al. 2020) and Bayesian inference analysis with MrBayes v. 3.2 (Ronquist et al. 2012). Model selection (Kalyaanamoorthy et al. 2017) implemented in IQ-TREE. Non-parametric bootstrapping (500 replicates) (Felsenstein 1985) was used to determine branch for the main tree (MAFFT aligned); 100 replicates for supplementary trees (and Bayesian analysis omitted for these trees).
Bayesian inference used variable rates and six rate categories. Two parallel runs of Markov chain Monte Carlo were performed for 3,000,000 generations, sampling every 1,000 generations. Estimated samples size, split frequencies, and stationarity were checked after each run. Post-analysis, 10% of generations were removed as a burn-in and posterior probabilities visualized in Figtree v1.1.4 (Rambaut 2009) and Canvas X Draw (Canvas GFX, Inc., Boston, MA, USA) were used to manipulate trees (e.g., collapse branches) for presentation.
The outgroups for all analyses were as in Buaya and Thines (2020): Hyphochytrium catenoides (AF163294, X80344), Developayella elegans (U37107), and an uncultured eukaryote (AB695482).
RESULTS AND DISCUSSION
To maximize the phylogenetic signal, we retained the maximum number of base pairs in the alignment so that after trimming, less than 20% of the partial SSU sequences were below 850 bp long. The different algorithms produced alignments of various lengths, with various numbers of variable (and parsimony-informative) sites and trees with different maximum-likelihoods (Table 1). All our analyses, as well as all previously published analyses (e.g., Buaya et al. 2019, 2023b, Buaya and Thines 2020) concur that the four supported clades containing holocarpic red algal parasites do not cluster together (Fig. 2), so that grouping these lineages into any single genus or single order (Fig. 2) is unwarranted. Even with the same data set, and different alignment methods, the relationships between these unsupported clades, as expected, vary (Supplementary Fig. S1).
Our investigation of the partial 18S sequences available for Pontisma lagenidioides and Sirolpidium bryopsidis (MK253530.2 and MW489460.1) further reveals that they are not identical over their entire length (818 bp) though conspecificity between both parasites was inferred because of “no genetic distance” between them (Buaya et al. 2023b). The identifications of the organisms concerned are also questionable: on the one hand, Sirolpidium bryopsidis had never been found before infecting Capsosiphon fulvescens. The latter host was collected in Iceland, whereas the type was described from Frederikshavn (Denmark), so that direct evidence is lacking that Sirolpidium bryopsidis is indeed the organism sequenced in Buaya et al. (2021). On the other hand, the host red alga depicted as the host for the epitype of Pontisma lagenidioides, is incompletely corticated (Buaya et al. 2019). Therefore, it is not C. rubrum as stated by the authors, and is in fact an unidentified Ceramium sp. These two examples illustrate that defining epitypes is only useful, to overcome the limitations of original taxonomic descriptions and to link them with molecular data, if utmost care is taken in the identification of host, choice of sampling location, and subsequent data analysis.
The novel parasite, Sirolpidium litorale, isolated from the green alga Urospora neglecta neither forms a clade with the known SSU sequence for S. bryopsidis (Buaya et al. 2023b, and our Fig. 1), nor seems to present typical features of this genus, such as fragmenting thalli (Petersen 1905, Sparrow 1960).
The absence of monophyly between the clades of red algal pathogens designated Pontisma or Sirolpidium by Buaya and colleagues, is now undisputable. Hence, the proposed circumscription of the genus Pontisma, or Sirolpidium, as a taxon encompassing pathogens of red algae, or for that matter any specific algal host, will have to be revised; instead, with the caveat highlighted above concerning the correct identification of its epitype, we propose that the genus name Pontisma should be restricted to Pontisma lagenidioides. We also reject the proposed synonymy between Pontisma and Sirolpidium for the time being and propose that the latter genus name remains restricted to its type, Sirolpidium bryopsidis, at least until (1) its presumed conspecificity with the parasite infecting Capsosiphon fulvescens is confirmed through the sequencing of endoparasites of Bryospis plumosa; and (2) a suitable epitype can be defined.
The Pontismatales, which has never been supported in any analyses should be disused. It was suggested that it could encompass red algal pathogens (Buaya and Thines 2020) but this was not correct, even in the Buaya and Thines (2020) analysis, where this was highlighted, it included diatom pathogens and subsequently found to also likely encompass brown algal pathogens (Buaya and Thines 2022, Buaya et al. 2023a). Our analyses clearly show that there are currently no groups that could be placed in a monophyletic supported order of biotrophic algal pathogens, as recently suggested (Buaya et al. 2023b). If it were confirmed as a monophyletic group, the concept of this order would require being broadened to accommodate far greater diversity of hosts (including brown algae and green algae), morphologies, and spore morphogenesis than is currently the case.
Over the last four years, two novel generic names, and the accompanying nomenclatural changes, Pontisma and Sirolpidium, have been proposed by Buaya et al. to designate species of parasites of marine red algae: Olpidiopsis bostrychiae, O. feldmanni, O. heterosiphoniae, O. muelleri, O. palmariae, O. porphyrae, and O. pyropiae. For the reasons stated above, it is hard to see any improvement over the original genus name Olpidiopsis, which at least has the merit of being widely used, especially by applied scientists and seaweed professionals from the industry and government bodies. Even when justifiable from a taxonomic point of view, name changes for ecologically significant and economically important species have deep consequences on society, potentially affecting livelihoods or conservation policies. Consequently, we argue in favor of retaining the names of these taxa until a robustly-supported nomenclatural revision can be performed.
Though we are aware of the challenges hindering the description and classification of holocarpic oomycetes, we feel that novel taxa should not be described without the provision of several congruent markers: indeed, relying on partial 18S sequences is insufficient. While many of these original descriptions are very insightful (e.g., Braun 1855, Sparrow 1960), attaching names to newly collected specimens, often on different hosts and different locations, from the original descriptions that were based on few characters (i.e., general similarity, sexual characters are missing in all red algal holocarpic pathogens) just brings more confusion, and retards naming and describing new taxa. We recommend that the nomenclature of supra-specific taxa should only be emended to produce a natural classification, when the corresponding taxa are precisely circumscribed with robust phylogenetic, morphological, and possibly ultrastructural or ecological criteria. Acquiring phylogenomic evidence on these taxa is most likely to be key to enable such a step.
CONCLUSION
The full diversity of biotropic oomycete pathogens is just now being explored. Our understanding of their relationships is still hampered by a lack of datasets that resolve their relationships. Premature nomenclatural changes based on unsupported relationships do not aid in furthering research, nor are they useful to interested parties. We propose that algal pathogens named “Olpidiopsis” remain that way until further data can be gathered to resolve relationships and more careful epitypification of old names be performed.