Abbreviations
AAPH
2,2′-azobis (2-amidinopropane) dihydrochloride
DCFH-DA
2,7-dichlorodihydrofluorescein diacetate
DMSO
dimethyl sulfoxide
DPPH
1,1-diphenyl-2-picryl-hydrazyl
DPPP
diphenyl-1-pyrenylphosphine
FBS
fetal bovine serum
HN
Haenam
JaH
Jangheung
JD
Jindo
JiH
Jinhae
MTT
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
PPP
polyphenols and proteins-rich extracts from P. yezoensis
ROS
reactive oxygen species
RPMI
Roswell Park Memorial Institute medium
SA
Sinan
WD
Wando
INTRODUCTION
Environmental stresses such as chemical agent and ultraviolet rays can disrupt the homeostasis between the antioxidant systems which plays an essential role in maintaining cellular balance (Sgherri et al. 2017). Reactive oxygen species (ROS) contain oxygen ions and peroxides and are key factors in antioxidant system (Choudhury et al. 2017). Excessed ROS production leads to the generation of superoxide anion radicals and hydrogen peroxide, and may underlie the pathology of diseases in human (Seifried et al. 2007). It has been proven that diseases such as inflammation and cancer often result from the oxidative damage of lipids, proteins, and even nucleic acids, leading to several physiological consequences (Diebold and Chandel 2016).
Over the last decades, polyphenols and proteins are reported to be effective nature antioxidants (Köksal et al. 2017, Ding et al. 2019). Polyphenols play as an antioxidant in human body; one of the main mechanisms of antioxidative activity in polyphenols is the H atom transfer, the single electron transfer and the metals chelation (Leopoldini et al. 2011). The mechanisms include the following two actions, one is non-specific actions based on chemical features which are more common to most polyphenols. Another one is specific mechanism based on particular structural and conformational characteristics which were identified by selected polyphenols (Fraga et al. 2010). This mechanism is associated with interactions of iron (Perron and Brumaghim 2009).
Brown algae are abundant in polyphenols while green and red algae have less. Bioactivities of Pyropia yezoensis are mostly related to proteins or peptides (Cian et al. 2015). In the study, the polyphenol contents of P. yezoensis were investigated in association with protein contents for antioxidant effects. In particular, polyphenols and proteins from marine resources have increased free radical scavenging activity and have been shown to inhibit the effects of lipid peroxidation (Cian et al. 2013). Compared to the synthetic drug for antioxidant, marine product becomes more flexible to obtain (Hamann 2007, Dai et al. 2020b). Thus, the exploration and screening the marine antioxidants is necessary to investigate.
P. yezoensis (Gim in Korean) is a popular seafood that is traditionally used as a marine vegetable along the East Asian coast (Kim et al. 2014b). It has been cultivated abundantly in Korea and is used to roll rice and other ingredients constituting the traditional Korean dish, gimbap (Aoki and Kamei 2006). Pyropia species are rich in different nutrients, including polysaccharides, polyphenols, sterols, glycoprotein, and lipids. They have several beneficial effects, such as antioxidant, anticancer, anti-inflammatory, and anti-fatigue activity and protective effects against ultraviolet rays-induced aging (Kwon and Nam 2006, Ryu et al. 2014, Lee et al. 2015, Choi et al. 2016, Hwang et al. 2018, Kim et al. 2018c). The antioxidant property of the acetone / water extract of Porphyra columbina was evaluated, and its polyphenol and peptide content showed high angiotensin-converting-enzyme inhibitory activity (Cian et al. 2013). We hypothesized that the polyphenols and protein-rich extracts from P. yezoensis (PPP) might exert antioxidant properties. In this study, we evaluate the protective properties of PPP against 2,2′-azobis (2-amidinopropane) dihydrochloride (AAPH)-induced lipid peroxidation in Vero cells and a zebrafish model, and find the best local strain of Korean from the six major locations for Gim aquaculture in Korea.
MATERIALS AND METHODS
Chemicals and reagents
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), 2,7-dichlorodihydrofluorescein diacetate (DCFH-DA), diphenyl-1-pyrenylphosphine (DPPP), dimethyl sulfoxide (DMSO), and Hoechst 33342 were obtained from commercial company (Sigma-Aldrich, St. Louis, MO, USA). All solvents used were of analytical grade.
Preparation of Pyropia yezoensis extracts
From 2015 to 2016, samples of P. yezoensis were collected from six areas along the south coast of the Korean Peninsula where major aquaculture farms are located; from Jinhae (JiH), Haenam (HN), Jangheung (JaH), Jindo (JD), Wando (WD), and Sinan (SA) (Fig. 1). Extracts containing the polyphenol- and protein-rich fraction from P. yezoensis followed the previous report (Kim et al. 2014b). Samples were cleaned with tap water to remove organic debris, salt, sand, and epiphytes and freeze-dried, maintained at −20°C. The sample of P. yezoensis from each area was ground to a powder; 3 g of each powder was shaken in 100 mL of 70% ethanol at 40°C for 24 h. The liquid layer was obtained via filtration, and then the filtrate was concentrated using an evaporator under vacuum pressure at 37°C. The extract after lyophilization was regarded as PPP. PPP was dissolved in DMSO, and then used for further experiments after adjusting the final concentration of DMSO in culture medium to <0.1%.
Chemical analysis
The approximate composition of the six samples from the different areas was evaluated according to the guidelines of the Association of Official Analytical Chemists methods. Therefore, the Kjeldahl method was applied to analyze the protein content, the Soxhlet method was applied to analyze the lipid content, and the muffle furnace (600°C for 6 h) was employed to analyze the ash content (Horwitz 1975). The determination of total polysaccharide and total polyphenol content were analyzed according to colorimetric standard methods (Dubois et al. 1956, Chandler and Dodds 1983).
Evaluation of antioxidant activities
The antioxidant activity of the extract was analyzed as a measurement of the 1,1-diphenyl-2-picryl-hydrazyl (DPPH) and alkyl free radical scavenging activities. The electron spin resonance spectrometer (JESFA200; Jeol, Tokyo, Japan) at 298 K was used for analysis. The DPPH and alkyl radical scavenging activities were analyzed according to the protocols followed by Nanjo and Hiramoto, respectively (Hiramoto et al. 1993, Nanjo et al. 1996).
Cell culture
Vero monkey kidney epithelial cell lines were purchased from Korean Cell Line Bank, Seoul, Korea. Roswell Park Memorial Institute medium (RPMI) and fetal bovine serum (FBS) were purchased from commercial company (Gibco Inc., Grand Island, NY, USA). Vero cells maintained at 37°C in a 5% CO2 incubator and cultured in RPMI medium supplemented with 10% (v/v) FBS, penicillin, and streptomycin (100 μg mL−1 each).
Antioxidant activity against AAPH-induced cell damage
MTT assay was employed to analyze the cell survival rate (%) after AAPH-induced cell death. AAPH is a promising water-soluble chemical for generate ROS (Mosmann 1983). The third passage Vero cells were seeded at a density of 1 × 105 cells mL−1 in 96-well culture plates. After seeding, the cells were treated with different concentrations (12.5, 25, and 50 μg mL−1) of PPPs. After 1 h incubation, cells were treated with AAPH (10 mM) for 24 h; untreated cells were used as control. The cell survival rate was evaluated using the MTT assay 24 h after sample treatment.
Nuclear staining with Hoechst 33342
To identify the protective effect of PPP on the AAPH-induced nuclear morphological changes in Vero cells, the Hoechst 33342 staining protocol was applied (Dai et al. 2019). Vero cells were seeded at a density of 1 × 105 cells mL−1 in 24-well culture plates. Cells were treated with different concentrations of PPPs from SA and JD for 1 h and spiked with 10 mM AAPH solution for 24 h. Then, Hoechst 33342 (stock, 10 mg mL−1) was added to each well at a final concentration of 10 μg mL−1 after 24-h incubation. Cells stained with 25 μL of Hoechst 33342 for 10 min and imaged using a CoolSNAP-Pro color digital fluorescent microscope (Olympus, Tokyo, Japan).
Origin and maintenance of parental zebrafish
Adult zebrafish were obtained from a commercial dealer (Jeju, Korea) and housed in a 3 L acrylic tank at 28.5°C with a 14 : 10-h light : dark cycle. The fish were fed with live brine shrimp three times daily. Embryos were collected by natural spawning, and the collection time of the embryos was controlled less to than 30 min.
Waterborne exposure of embryos to PPP and AAPH
The embryos (n = 15) were randomly transferred to each well of 24-well plates and grown in embryo medium between 7 to 9 h post-fertilization (hpf). Then, different concentrations (12.5, 25, and 50 μg mL−1) of PPP from SA were added to each well. After incubation for 1 h, 25 μL of 15 mM AAPH solution was added to the wells up to 24 hpf. The embryo medium was substituted with fresh medium.
Measurement of AAPH-induced intracellular ROS generation and image analysis
DCFH-DA, an oxidation-sensitive fluorescent probe dye, was used to determine the generation of ROS in embryos. At 7–9 hpf, the embryos were treated with different concentrations (12.5, 25, and 50 μg mL−1) of PPP and incubated. One hour later, 25 μL of 15 mM AAPH solution was added to the plate. After 24 h, the embryo medium was substituted with fresh medium. The embryos were transferred into a 96-well plate, which was subsequently spiked with DCFH-DA solution (20 μg mL−1) and incubated in the dark at 28.5°C for 1 h. Then, the embryo medium was rinsed off, and embryos were anesthetized before observation. A spectrofluorometer (Perkin-Elmer LS-5B, Vienna, Austria) was used to calculate the fluorescence intensity of each embryo, and the CoolSNAP-Pro color digital fluorescent microscope (Olympus) was used to quantify and image the stained embryos (Dai et al. 2020a).
Measurement of oxidative stress-induced cell death in zebrafish embryos
Acridine orange preferentially stains necrotic or late apoptotic cells because acridine orange can disturb plasma membrane permeability. At 4 hpf, the embryos were exposed to different concentrations (12.5, 25, and 50 μg mL−1) of PPP from SA. One hour later, 25 μL of 15 mM AAPH solution was added to each plate. The embryos were maintained until 2 dpf, and the medium was replaced with fresh medium every 24 h. The treated embryos were transferred into a 96-well plate, and acridine orange stain was added at a concentration of 7 μg mL−1; the plate was incubated for 30 min in the dark at 28.5°C. Before visualization, the embryo medium was replaced with fresh medium and the embryos were anesthetized. The fluorescence intensity and images of embryo were quantified and observed, followed by the protocol of “measurement of AAPH-induced intracellular ROS generation and image analysis.”
Statistical analysis
All assays involved three independent experiments. Values are expressed as the mean ± standard error (SE), and one-way ANOVA (SPSS statistical software; IBM Corp., Armonk, NY, USA) was used to analyze the mean values. Student’s t-test was used for the analysis of the means of parameters showing significant difference.
RESULTS
Chemical composition of PPP obtained from six locations
P. yezoensis samples were collected from six different locations across South Korea, and their chemical compositions were determined. As shown in Table 1, the extract from samples obtained in WD showed the highest ash content, followed by that from JD. The highest polyphenol and carbohydrate content were observed in the samples from SA and JD. The samples from SA and JD had the highest total sterol content, followed by the others. The highest protein content was observed in samples from SA and HN. The highest extract yield was shown by the sample from JD at 20 ± 0.2% followed by that from WD at 16.2 ± 0.3% yield. The moisture content slightly varied among the samples. Interestingly, SA and JD are both located towards the western coast of Korea and, among the six samples, the samples from these areas showed relatively high polyphenol, carbohydrate, total sterol, and protein content. Factors such as sample condition, water temperature, and the intensity of sunlight may have an influence on the content of polyphenols and other active compounds, which could be investigated further in future.
Radical scavenging activities of PPP
PPP from SA showed the highest DPPH and alkyl radical scavenging activities, with IC50 values of 0.05 ± 0.01 and 0.19 ± 0.02 μg mL−1, respectively (Table 2). PPP from HN showed the second highest alkyl radical scavenging activities with IC50 values of 0.16 ± 0.03 μg mL−1 among these six PPPs.
Antioxidant activity of PPP against AAPH-stimulated damage in Vero cells
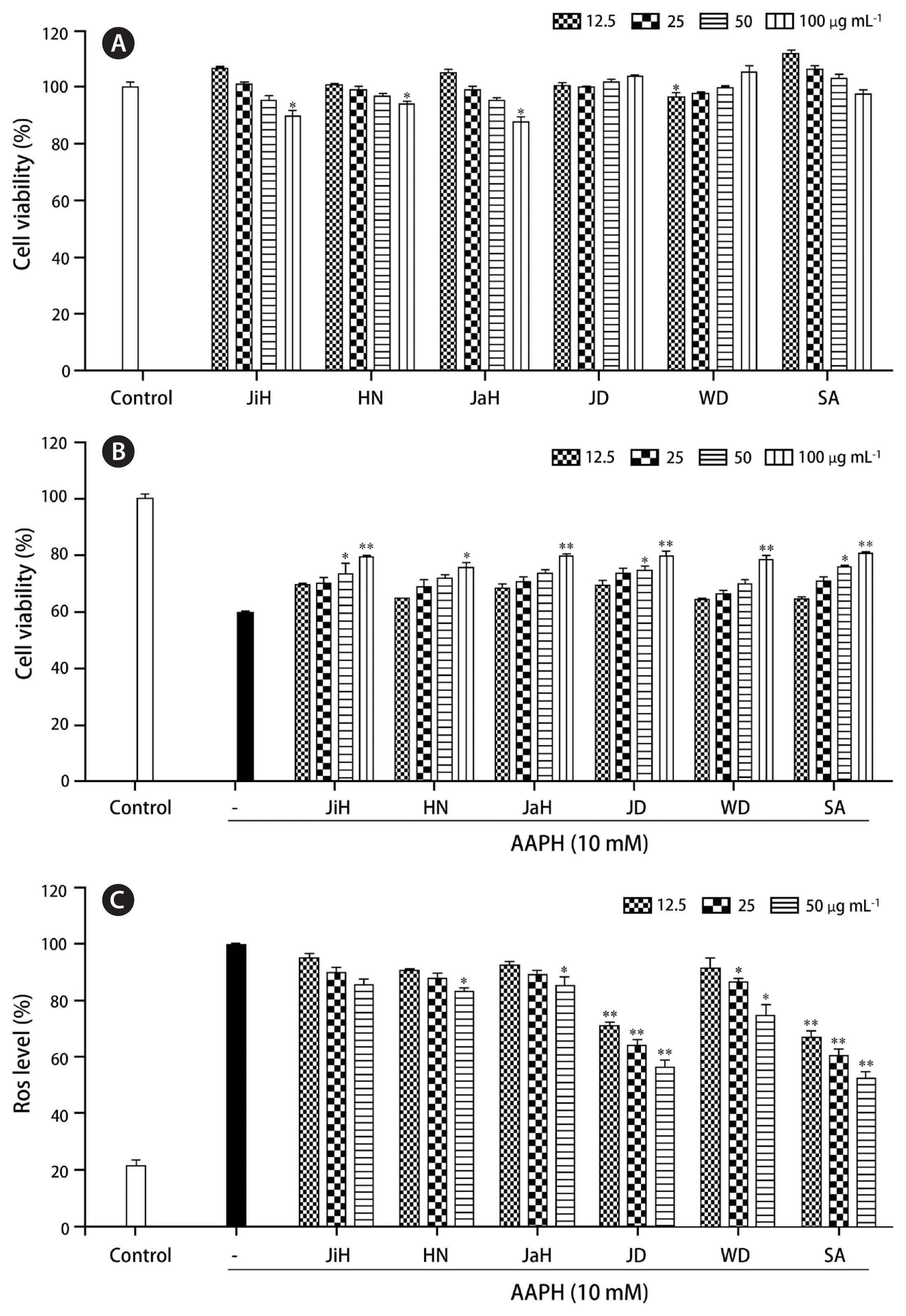
The cytotoxicity of PPP from the P. yezoensis samples collected in the six different areas was determined on Vero cells using the MTT assay. Vero cells are one of the most commonly and promising used mammalian kidney cells, which become the best cell line for antioxidant effect in vitro (Ammerman et al. 2008). No cytotoxic effects of the six P. yezoensis were observed (Fig. 2A). Subsequently, we analyzed the protective effects of the six P. yezoensis samples against AAPH-induced cell damage using the MTT assay. Upon exposure of AAPH to Vero cells, pretreated with 12.5 to 100 μg mL−1 concentrations of PPP, the cell viabilities indicated higher than AAPH-treated group (Fig. 2B). This result demonstrated PPPs have the protective effect in AAPH-treated Vero cells. ROS production was assessed in the Vero cells pretreated with PPPs followed by exposed to AAPH. Significant decreases in the ROS level were observed in the treated samples with JD and SA compared to the untreated AAPH-stimulated control; this indicated the strong scavenging properties of PPP (Fig. 2C).
Apoptotic body formation in Vero cells upon treatment with PPPs from SA and JD
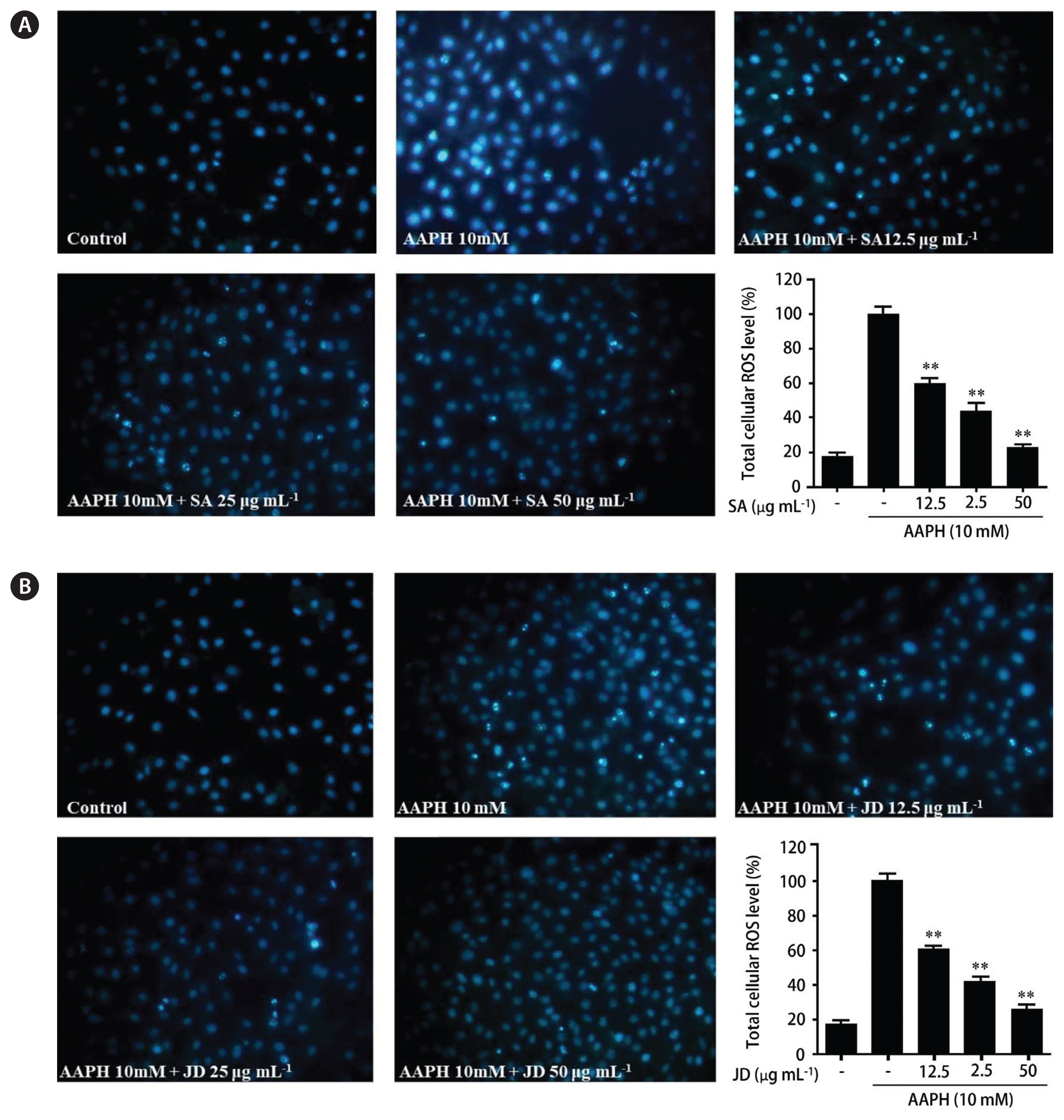
Since PPPs from SA and JD significantly decreased ROS production amongst the six samples, SA and JD were selected for further investigation. The nuclear morphology of Vero cells, pretreated with PPPs and exposed to AAPH, was examined by Hoechst 33342 staining followed by fluorescence microscopy. Apoptotic body formation was seen in the AAPH-treated cells; these results are shown in Fig. 3. Cells pretreated with SA and JD showed decreases in the amount of apoptotic bodies in a dose-dependent manner.
Toxicity of AAPH and PPP treatment in zebrafish embryos
PPP from SA showed minimal toxicity compared to the control (data not shown). Therefore, the viability of zebrafish embryos, treated with PPP from SA and exposed to AAPH, was evaluated; these results are shown in Fig. 4. The AAPH-induced embryos showed lower viability compared to the control, whereas the embryos pretreated with SA showed a survival rate of 50–80% post-AAPH treatment.
Protective effect of PPP from SA against AAPH-induced oxidative stress and cell death in zebrafish
The fluorescence intensity of acridine orange, a nucleic acid-specific metachromatic dye, was used to determine cell death and evaluate the protective effect of PPP from SA against AAPH-stimulated cell death in the zebrafish model. Cell death in AAPH-treated zebrafish embryos was 232.9% increase compared than untreated embryos; however, the pretreatment with different concentrations of SA significantly decreased cell deaths (Fig. 5).
The antioxidative activity of SA on ROS production in AAPH-induced zebrafish embryos was observed using the DCFH-DA detection assay. The embryos pretreated with PPP from SA showed a strong inhibitory effect on ROS production. The control group (untreated zebrafish embryos) exhibited low fluorescence intensity, whereas AAPH treatment led to a significant increase in fluorescence. This suggested that ROS production in the zebrafish embryos increased upon stimulation with AAPH.
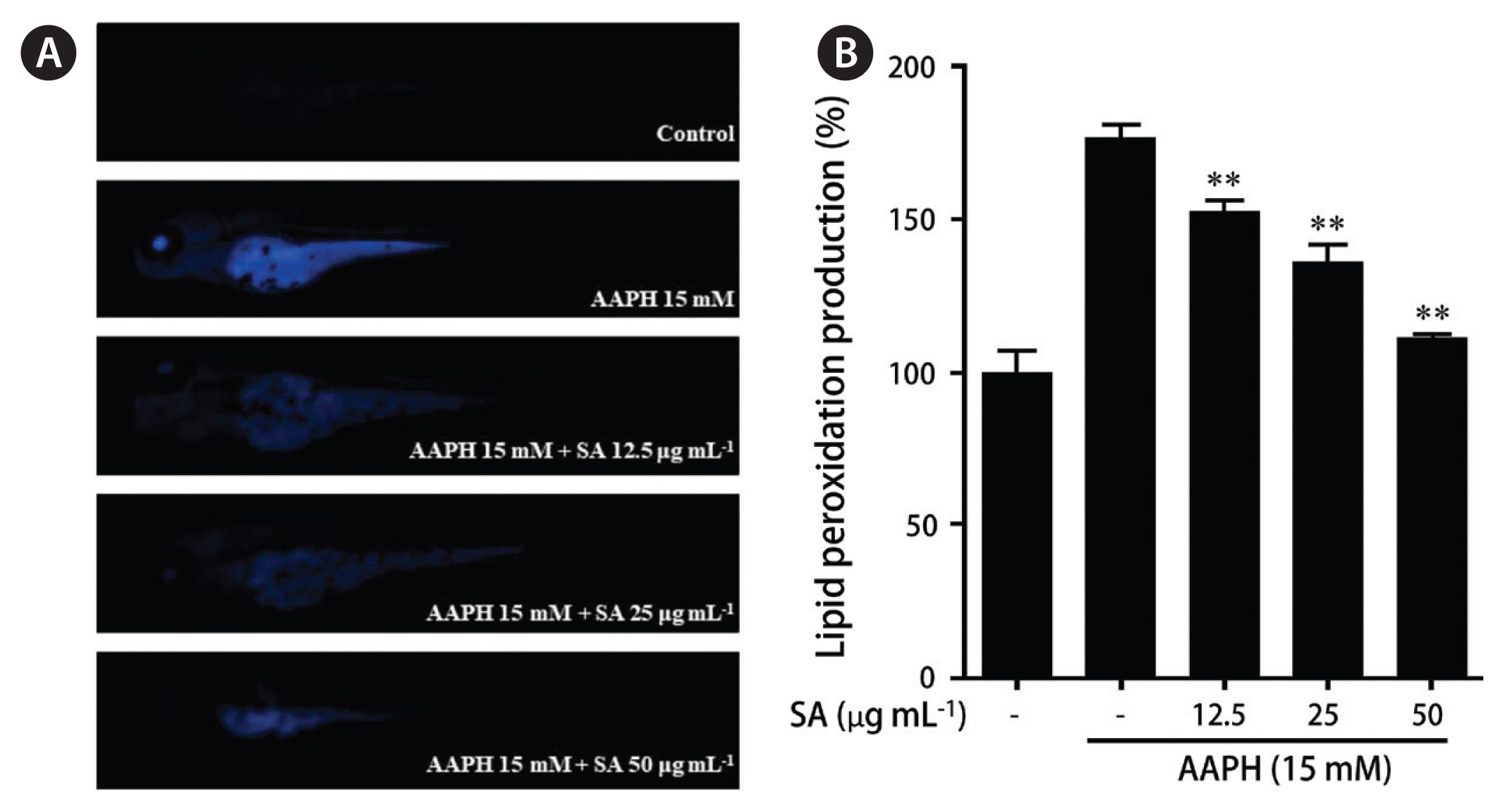
DPPP is widely used to evaluate lipid peroxidation in zebrafish. The untreated embryos showed lower fluorescence, whereas the treatment with AAPH resulted in higher fluorescence than the control, indicating that AAPH treatment induced lipid peroxidation. However, the pretreatment with SA decreased the fluorescence intensity of AAPH-induced embryos suggesting that the treatment affords protection against lipid peroxidation (Fig. 6). Collectively, these results demonstrate that PPP from SA could be a potential natural antioxidant.
DISCUSSION
The environmental factors affect the quality of seaweeds in various ways. The antioxidant and anti-inflammatory activities of methanol extracts of were evaluated in brown seaweed, Undaria pinnatifida using local strains collected from southeastern sea shores in Korea (Hwang et al. 2014). The different antioxidant and anti-inflammatory activities of the seaweeds in the three areas indicated differences chemical compositions and different inhabited areas (Sanjeewa et al. 2019). The climates of the eastern and western parts of Korean Peninsula are influenced by ocean currents (Tanaka et al. 2012, Kim et al. 2018b). Therefore, we believe that locations strongly influence the quality of Gim. In this study, we collected P. yezoensis from six regions where major Pyropia aquafarms locates screened their antioxidant effects both in vitro and in vivo.
Environmental stressors such as cold shock and UV-rays as well as biotic stressors like viral infection cause the accumulation of ROS, leading to lipid peroxidation, cell damage, and death. AAPH is a water-soluble compound that has long been known to be a free radical generator (Niki 1990). In this study, we used AAPH to induce oxidative stress in Vero cells and zebrafish embryos. Typically polyphenols are rich in brown seaweed (Namvar et al. 2013), and they exhibit strong antioxidant activity against DPPH, hydroxyl, and peroxyl radicals (Kang et al. 2012). There have been very few studies on the extracts of red seaweed and their bioactivity and polyphenol content, particularly because of the lower levels of these compounds in these seaweeds than brown seaweeds. Hence, the content and antioxidant effects of polyphenols extracted from the red seaweed P. yezoensis, collected from different areas, were studied. Our results demonstrated that the content of polyphenols and protein in the 70% ethanol extract of P. yezoensis was relatively higher than that of other compounds. Among the samples collected from the six areas, those from HN and SA exhibited the strongest antioxidant effects against alkyl radicals, with recorded IC50 values of 0.16 ± 0.03 and 0.19 ± 0.02 mg mL−1, respectively. SA and JD exhibited the strongest antioxidant effects against DPPH radicals, with recorded IC50 values of 0.05 ± 0.01 and 0.26 ± 0.02 mg mL−1, respectively. Additionally, the total sterol contents of the extracts from SA and JD were higher than those of the other samples, at 0.34 ± 0.01 and 0.33 ± 0.01%, respectively. The sterol fraction from Porphyra dentata has been reported to possess anticancer effects in vitro and in vivo (Kazłowska et al. 2013). Hence, we hypothesize that the sterol content in P. yezoensis may also contribute to its antioxidant activity. The antioxidant effect of PPP against AAPH-induced cell death was evaluated using Vero cells. The PPPs from SA and JD were found to decrease the cell death and ROS production in a dose-dependent manner in AAPH-induced Vero cells; this clearly exhibited their protective effects against oxidative damage. Furthermore, treatment with PPPs from these samples also decreased the formation of apoptotic bodies in AAPH-stimulated Vero cells in a dose-dependent manner. These results confirm our hypothesis that PPPs from SA and JD show strong antioxidant properties. All the results confirmed our hypothesis that the antioxidant effect of SA and JD in Vero cells was exerted via anti-apoptosis manner, which was mediated by a signaling cascade.
In pharmacological studies, the zebrafish, as an in vivo model, has unique advantages, such as large clutch size, transparent embryos, low cost, and easy handling. Hence, it is routinely used for screening in predictive toxicology (Howe et al. 2013). In the zebrafish model, lipid peroxidation was evaluated using a fluorescent probe DPPP, which shows diminished fluorescence under peroxidized conditions. Cell death was observed in zebrafish embryos by acridine orange staining, a nucleic acid-specific metachromatic dye (Kim et al. 2014a). We have also validated our hypothesis using zebrafish embryos as an in vivo model system. The PPP from SA inhibited the AAPH-simulated cell death and ROS generation in zebrafish. Lipid peroxidation is a well-characterized toxicological phenomenon that is most commonly initiated by ROS. The free radical generator and AAPH can stimulate lipid peroxidation in cells leading to cell death (Yokozawa et al. 1997). Recent studies reported that stimulator-induced lipid peroxidation in zebrafish embryos was decreased by the polyphenol-rich fraction obtained from seaweeds (Kim et al. 2014c). It was proven that PPP from Korean Gim (SA) has a strong protective effect against lipid peroxidation and cell death in AAPH-treated zebrafish embryos. Lipid peroxidation is a chain reaction in which the reaction of lipids with a free radical generates lipid hydroperoxide and peroxyl-fatty acid radicals; these subsequently attack different biological molecules resulting in a sustained reaction that can induce physiochemical alterations, ultimately leading to cell death (Miki et al. 1987). Hence, PPP from SA exerted protective effects against the damage caused by lipid peroxidation.
In this study, subsequently, the free radical scavenging and protective activities of PPP on AAPH-stimulated oxidative stress using cells and zebrafish embryos were evaluated. PPP extracts from P. yezoensis obtained from six locations were analyzed. Of these, SA and JD contained relatively higher contents of polyphenols and protein and showed the strongest radical scavenging activity against AAPH and the protective activity of PPP against AAPH-simulated intracellular ROS production and cell death of Vero cells. Finally, it was found that Korean Gim from SA and JD strongly inhibited AAPH-simulated cell death, ROS generation, and lipid peroxidation in zebrafish.
Thus, our studies using Vero cells and zebrafish embryos have demonstrated the antioxidant, free radical scavenging, and protective activities of the polyphenol and protein-rich extract of P. yezoensis. The product from SA and JD, which located southwestern coast of Korea, showed the strongest antioxidant activity partly because of the environmental factors. These results suggest that the geographical location of algae could be a major determinant in its chemical content and activities even when the same species was used for culture.