INTRODUCTION
Sterols are one of the important component in the membrane of all eukaryotic cells (Volkman et al. 1998). Moreover, cholesterols are major sterols of vertebrate organisms and mixture of various phytosterols containing in photosynthetic organisms (Lopes et al. 2013). Farnesyl diphosphate convert into squalene and this step activate the isoprenoid pathway which capable of producing sterols in biological systems (Hartmann 1998). Sterols located in microalgal cell membranes act as biomarkers for identifying organic matters in their surroundings and acting as an essential component for cell proliferation (stigmasterol) and signal transduction in microalgae (Hartmann 1998, Volkman et al. 1998). Moreover, phytosterols separated from marine microalgae found to possess interesting bioactive properties, including anti-inflammatory, anti-cancer, antioxidant, antidiabetic, and neuromodulatory activity (Yasukawa et al. 1996, Francavilla et al. 2012, Kim et al. 2014, Luo et al. 2015).
Cancer developed in multicellular organs when the balance between the cell generation and cell death is altered (Cotter 2009). Cancer is considered as one of the major threats to human thus been made a high level of suffering and economic losses to entire humans. Due to the high level of impact, considerable attention has been made to eliminate cancer (Jayaprakasam et al. 2003). Some reports suggest that there has been a positive relationship between inflammation and tumor progression with anticancer compounds (Coussens and Werb 2002). Most of the anticancer drugs developed to treat cancer have been shown to induce apoptosis in cancer cells. Moreover, those anticancer drugs induce apoptosis cell death via, inducing the endonucleolytic cleavage of DNA and changes in chromatin condensation of cancer cells (Hickman 1992). Furthermore, phytosterols separated from microalgae have identified as an excellent source for develop chemopreventive drugs for cancer diseases. Moreover, a number of studies have reported phytosterol separated from microalgae induce apoptosis cell death in cancer cells (Luo et al. 2015).
Inflammation can define as the normal protective response of living tissue to injury or infection. However, an inflammatory response is a series of local cellular and vascular responses that immediately occurs following tissue damage as a result of either, physiological or pathological processes and act as a protective mechanism of organisms to remove the noxious stimuli from the tissues and to initiate the healing process (Calder 2006, Medzhitov 2008, Huang et al. 2012).
During inflammation, inflammated cells start to produce pro-inflammatory cytokines as well as stimulate the expression of nitric oxide synthase isoform and an inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2). Furthermore, expression of iNOS led to produce tiny, reactive radical gas known as nitric oxide (NO) in phagocytes including macrophages and dendritic cells and COX-2 led to produce protaglandin E2 (PGE2) in lipopolysaccharide (LPS)-stimulated RAW cells (Nathan 1992, Lowenstein et al. 1993, Gordon 2002, Lee et al. 2002).
Nannochloropsis species belong to the family eustigmatophyte and are photosynthetic eukaryotic unicellular microorganism’s habitats in both fresh and sea water environments. Moreover, eustigmatophyte microalgae species are popular as a good source of phytosterols (Sukenik et al. 1993, Converti et al. 2009). However, knowledge on bioactive properties of Nannochloropsis species are limited and very low. The aim of this study was to investigate the effects of partially purified sterol-rich hexane fraction of marine microalga Nannochloropsis oculata on anti-cancer and anti-inflammatory activities.
MATERIALS AND METHODS
Chemicals and reagents
Five cancer cell lines including human promyelocytic leukemia cell line (HL-60), human colon adenocarcinoma cell line (SW-480), adenocarcinomic human alveolar basal epithelial cell line (A549), hepatocellular carcinoma cell line (Hep-3B), and colorectal carcinoma cell line (HCT-116) were used to evaluate anti-cancer activity and murine macrophage cell line (RAW 264.7) was used to measure in vitro NO production and African green monkey kidney (Vero) cell line was used to evaluate the cytotoxic effect of tested concentrations. All cell lines were purchased from the Korean Cell Line Bank (Seoul, Korea). Roswell Park Memorial Institute (RPMI-1640) medium, Dulbecco’s modified Eagle’s medium (DMEM), penicillin-streptomycin and fetal bovine serum were purchased from Gibco/BRL (Burlington, ON, Canada). 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) and dimethyl sulfoxide (DMSO) were purchased from Sigma Aldrich (St. Louis, MO, USA). All other chemicals and reagents used in these experiments were of analytical grade.
Culture of marine microalga
Marine microalga N. oculata was provided by the Korea Marine Microalgae Culture Center. The alga was pre-cultivated in 5 L glass vessels (medium 4 L) and then inoculated in 30 L plastic cylinders at 20°C, although air was supplied continuously at 5 L min−1 by air-lift. Sixty watt fluorescent lamps were used to provide light energy at an intensity of 34 μmol photons m−2 s−1 throughout the growing period. The microalga was cultured in Conway medium as previously described by Walne (1966). In brief, medium was prepared from filter-sterilized seawater and the culture was continuously active during the 8 to 10 days after inoculation. The cells were flocculated with 200 ppm Al2(SO4)3 (v/v) (Magicpool-99; Ilshin Biochemical, Hanam, Korea), and then recovered with centrifugation at 2,000 rpm using a basket centrifuge (Hanseong Co., Ansan, Korea). The harvested N. oculata biomass was frozen at −80°C and preserved until freeze dry.
Determination of proximate chemical compositions of Nannochloropsis oculata
Proximate composition of moisture, protein and ash contents were determined using standard Association of Official Analytical Chemists (AOAC, 1998) methods (Cunniff 1998). Crude protein was measured by calculating a conversion factor of 6.25. Crude lipid content was measured by using Soxhlet method with diethyl ether solvent (Soxtec 2050; FOSS Analytical, Hillerød, Denmark) and crude ash was determined by incineration of the sample at 600°C in the muffle furnace (B180; Nabertherm GmbH, Lilienthal, Germany). Moisture content was determined by the oven-drying method at 105°C in the moisture analyzer (mb45; OHAUS, Nänikon, Switzerland).
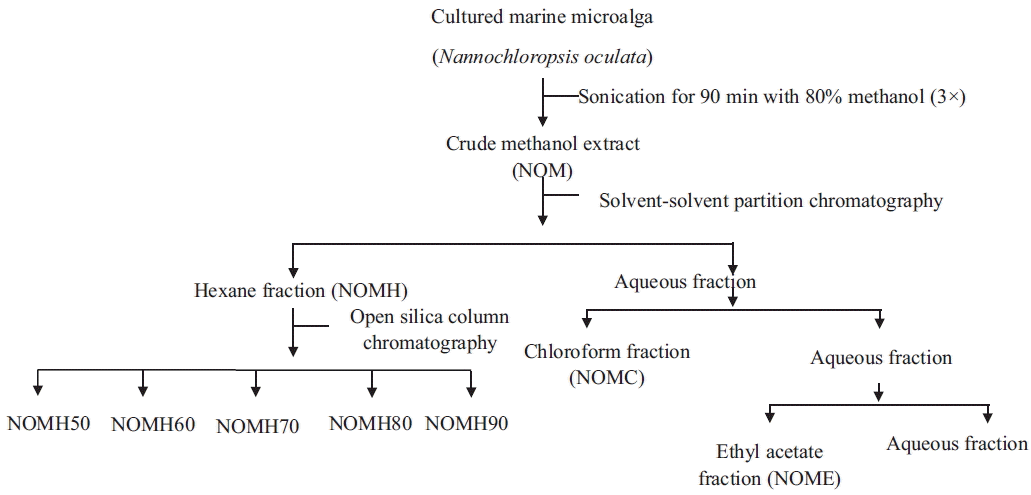
Sample preparation and fractionation
The freeze dried N. oculata sample (10 g) was extracted three times at less than 40°C for 90 min with 80% methanol solution (1.0 L) using an ultrasonic bath (JAC 2010; Kodo Technical Research Co. Ltd., Hwaseong, Korea). The liquid layer was obtained via vacuum filtration, and the filtrates were concentrated using rotary evaporator followed by freeze dryer. The freeze dried powder was considered as the crude methanolic extract of N. oculata (NOM).
Then the NOM powder was dissolved in distilled water and partitioned according to the polarity using n-hexane (NOMH), chloroform (NOMC), and ethyl acetate (NOME), respectively. The resulting solvent fractions were concentrated using rotary evaporator and freeze-dried. Further fractionation of an active fraction (NOMH) was conducted using an open silica column by hexane / ethyl acetate step-gradient elution with successive ratios of 90 : 10 (NOMH90), 80 : 20 (NOMH80), 70 : 30 (NOMH70), 60 : 40 (NOMH60), and 50 : 50 (NOMH50). Each fraction was evaporated by using a rotary evaporator (Fig. 1).
Total sterol content measurement of Nannochloropsis oculata extracts
Total sterols contents of N. oculata extract and its fractions ware assessed by cholesterol quantification using Liebermann-Buchard reaction with some slight modifications (Huang et al. 1961, Costa et al. 2013). Briefly, 150 μL of Liebermann-Burchard reagent (Burke et al. 1974) were mixed with 5 μL of each sample (in a 96-well plate) followed by incubation at 37°C for 30 min. Absorbance was measured at 625 nm in a spectrophotometer and the sterol concentration was determined from a standard curve for cholesterol content.
Cell experiments
Vero cells and all cancer cell lines were grown in RPMI medium, while RAW 264.7 cells were cultured in a DMEM medium. Both culture mediums were supplemented with 10% heat-inactivated FBS and 1% streptomycin (100 μg mL−1), and penicillin (100 unit mL−1). The cells were maintained and incubated in modified atmospheric condition which contained 5% CO2 and at 37°C (ambient temperature). Cells were sub-cultured within 2 days intervals and exponential phase cells were used for experiments.
Determination of cytotoxicity on Vero cells and anti-cancer activities of extracts
MTT assay, is a test of metabolic competence predicated on the assessment of mitochondrial performance, which depends on the reduction of the yellow color tetrazolium salt MTT into its blue-black formazan crystals via mitochondrial succinate dehydrogenase in viable cells (Pieters et al. 1988). Cell cytotoxicity of all cell lines were estimated via a colorimetric MTT assay as described by Mosmann (1983), with slight modifications. Briefly, 190 μL of cells were seeded in a 96 well plates with 1 × 105 cells mL−1 concentration and plates were incubated for 24 h, and then treated with 10 μL from measured concentration (6.25, 12.5, or 25 μg mL−1) of NOMH fractions and again incubated for 24 h. Ten microliter of MTT reagent (2 mg mL−1 in phosphate buffered saline [PBS]) was added to each well and aspirate the medium after 3 h of incubation and formazan crystals were dissolved in DMSO, and then the amount of blue-black formazan was measured using enzyme-linked immunosorbent assay (ELISA) reader at 540 nm.
Determination of NO production
NO production was assayed using a method described by Leiro et al. (2002) with some modifications. On the basis of nitrate production in RAW 264.7 culture supernatants, measured with Griess solution. Briefly, 450 μL of cell suspensions (1 × 105 cells mL−1) were incubated on 24 well plates for 24 h and then treated with 25 μL of measured concentrations of NOMH fractions to each well and further incubated for 1 h and treated with 25 μL of LPS and further incubated for 24 h. Then 100 μL cell suspensions were mixed with equal volume of Griess reagent. After 10-min incubation, the absorbance was read with an ELISA reader at 540 nm.
Western blot analysis
Various concentrations of (3.12, 6.25, and 12.5 μg mL−1) the active fraction were used to determine anti-inflammatory protein expression on RAW 264.7 cells. Proteins were extracted with PRO-PREP protein extraction kit (iNtRON Biotechnology, Seongnam, Korea). Briefly cell lysates were centrifuged 16,000 rpm at 4°C for 20 min to collect supernatants. The protein concentration of supernatants were determined by using Bio-Rad protein assay kit (Bio-Rad, Hercules, CA, USA). The cytoplasmic proteins (40 μg) were electro-transferred onto a nitrocellulose membrane following separation on a 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis under denaturing conditions. After blocking with 5% nonfat milk for 1 h, the blots were separately incubated with specific primary rabbit polyclonal anti-rabbit iNOS or mouse monoclonal anti-mouse COX-2 or β-actin (Cell Signaling Technology, Beverly, MA, USA) antibody for 60 min and washed twice with Tween 20/Tris-buffered saline (TTBS). Then, the blots were incubated with horseradish peroxidase–conjugated anti-mouse or anti-rabbit IgG for 45 min followed by visualized using an enhanced chemiluminescence (ECL) reagents (ECL; Amersham, Arlington Heights, IL, USA) onto a nitrocellulose membrane.
Nuclear staining with Hoechst 33342
HL-60 cells were cultured in 24-well plates at a concentration of 1 × 105 cells mL−1 and cells were incubated at 37°C for 24 h. The cells were then treated with various concentrations of NOMH90 (6.25, 12.5, and 25 μg mL−1) and incubated for an additional 24 h again at 37°C. Then, Hoechst 33342, a DNA-specific fluorescent dye was added to the culture media at a final concentration of 10 μg mL−1 and the plates were incubated for an additional 10 min at 37°C. The stained cells were then observed under a fluorescence microscope equipped with a CoolSNAP-Pro color digital camera in order to determine the degree of nuclear condensation.
Cell cycle analysis
HL-60 cells at a density of 1 × 105 cells mL−1 were treated with various concentration of NOMH90 (6.25, 12.5 and 25 μg mL−1) or the solvent alone. After 24 hours, the cells were harvested by centrifugation, washed twice with PBS and suspended in 1 mL 70% ethanol solution at 4°C for 30 min. After 30-min incubation cells were centrifuged, and the cell pellets were stained with 4′,6-diamidino-2-phenolindole (DAPI). DNA content was then analyzed with a FACSCalibur flow cytometer (Becton Dickinson, San Jose, CA, USA).
Statistical analysis
All the data were expressed as mean ± standard deviation of three determinations. Analysis of variance was carried out for the collected data using the Statistical Package for Social Science (IBM SPSS Statistics for Windows, version 20.0; IBM Corp., Armonk, NY, USA) statistical analytical package. Mean values of each experiment were compared using one-way analysis of variance. The Duncan’s multiple range test was used for the mean separation. A p-values less than 0.05 was considered significant.
RESULTS
Proximate chemical composition and sterol contents of the microalgal samples
The proximate composition and total sterol content of N. oculata were investigated. Total protein content was recorded as 30.52% dry weight basis (DW). Ash and lipid contents were respectively recorded as 30.64% and 7.99% DW (Table 1). Sterol contents of the algal extract and its fractions were measured using Liebermann-Buchard reaction (Table 2). According to the results, sterol content of NOM was recorded as 19.38% and phytosterols contents of its solvent fractions (NOMH, NOMC, and NOME) ranged between 32.35 to 11.36% and the highest was recorded from NOMH fraction. As well as bioactivities of NOM fractions were also increased with sterols contents. Hence, study was continued only with NOMH fraction. Furthermore, five fractions eluted from NOMH fraction by using open silica column were also measured for the sterol content. The sterol contents of the five NOMH fractions were ranged between 6.34 to 63.59% and the highest was recorded from the NOMH90 while the lowest from NOMH50 fraction.
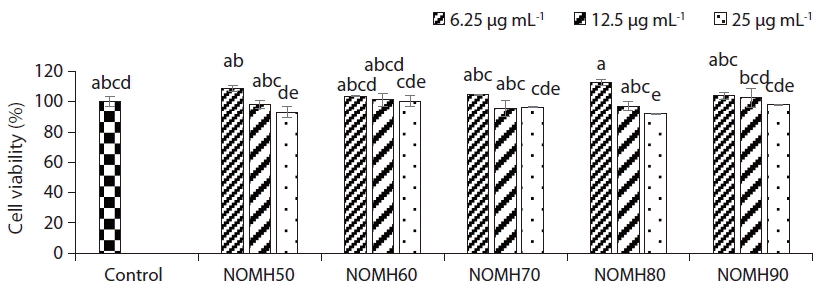
Cytotoxicity assay
The cytotoxicity study was carried out for each NOMH fraction to measure optimum sample concentration for the growth of Vero cells. The effects of NOMH fractions on the cell viability were determined with the MTT assay (Fig. 2). According to the cell viability results, all the tested fractions did not exhibit cytotoxic effect on the Vero cells, except 25 μg mL−1 concentration of NOMH80 (92% cell viability), which showed a significant cytotoxicity on the cells compared to the control (Fig. 2). Therefore, it revealed that all the fractions tested concentrations are not cytotoxic to the cells.
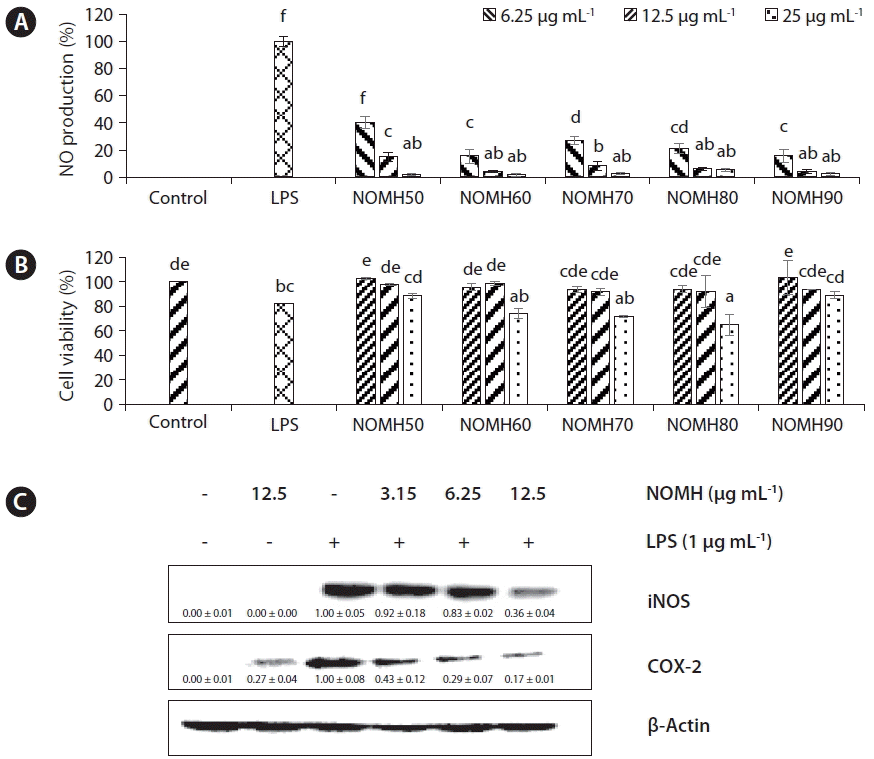
Evaluation of anti-inflammatory activity of NOMH fractions on RAW 264.7 macrophages cells
To investigate the anti-inflammatory effects of the five NOMH fractions, the inhibitory effects on NO production were examined in LPS activated RAW 264.7 macrophage cells. All the five fractions inhibited NO production on macrophages cells in a dose-dependent manner, compared to the LPS treated cells as shown in Fig. 3A.
RAW 264.7 cell viability in the presence of NOMH fractions are shown in Fig. 3B. According to the results NOMH50 and NOMH90 fractions were not significantly affected on cell viability of RAW 264.7 cell. However, NOMH60, NOMH70, and NOMH80 fractions at the 25 μg mL−1 exhibited significant cytotoxicity. According to the results, NOMH90 fraction has been subjected to Western blot analysis in which NOMH90 fraction down-regulated the iNOS and COX-2 protein expression in a dose-dependent manner as shown in Fig. 3.
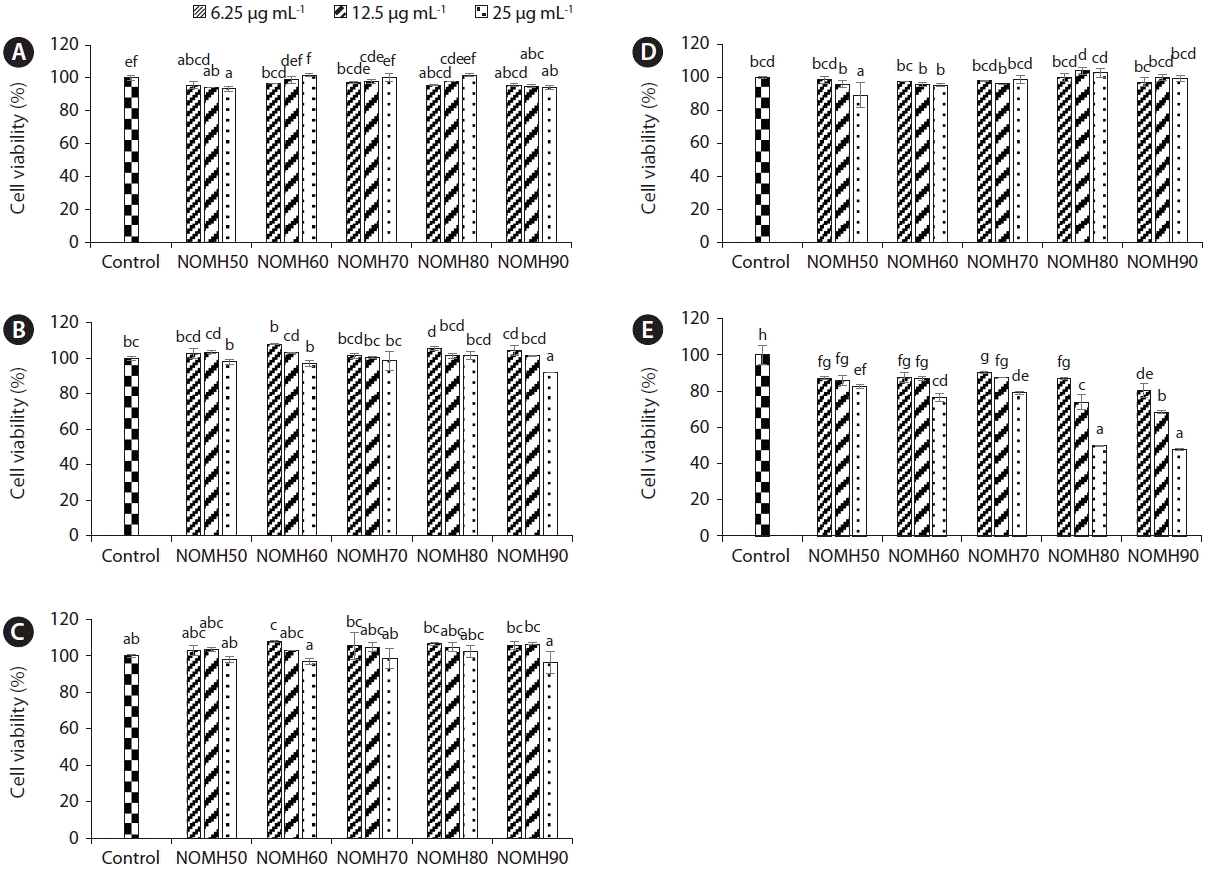
Anti-proliferative effect of NOMH fractions against the growth of selected five cancer cell lines
Cell proliferation is a crucial marker for the promotion and progression of carcinogenesis. In this study we measured cell viability percentage of five respective cancer cell lines HL-60, A-549, HEP-3B, HCT-116, and SW-480 incubated with different concentrations of NOMH (Fig. 4). According to the results NOMH80 and NOMH90 showed strong anticancer effect against HL-60 cells in dose-dependent manner. Moreover, according to the results IC50 values of NOMH80 and NOMH90 on HL-60 cells were 24.81 ± 0.34 and 23.58 ± 0.09 μg mL−1, respectively.
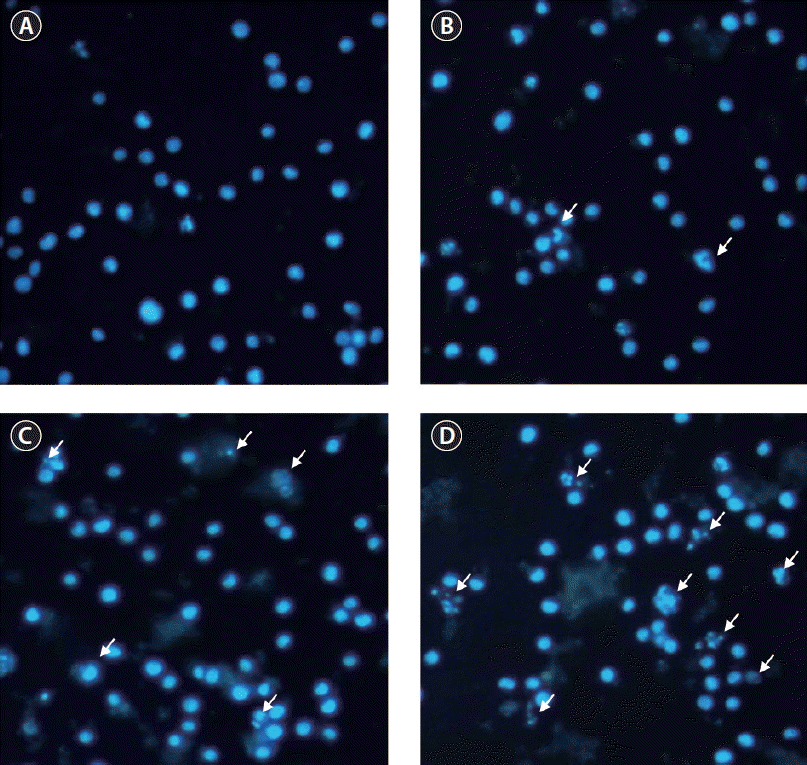
Apoptosis morphological changes observed under fluorescent microscopy
Apoptosis can be defined as breakdown of cell DNA into the small size basal points (180–200) or 50–300 kb ranged large DNA fragments (Lizard et al. 1995), hence cells with homogeneously stained nuclei were considered viable, whereas the presence of chromatin condensation or fragmentation was indicative of apoptosis. Fig. 5 noticed that NOMH90 dose-dependently increased the proportion of apoptosis body formations in HL-60 cells.
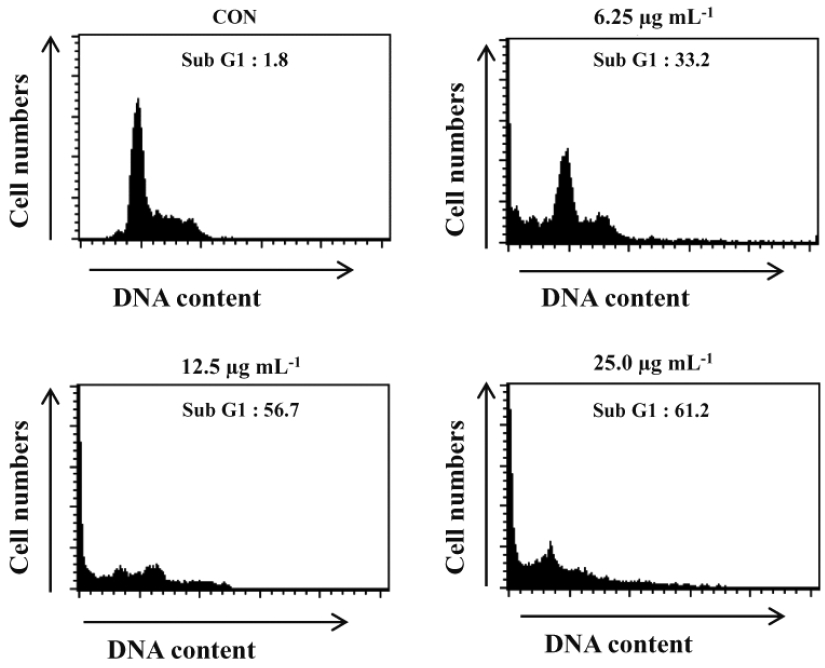
Induction effect of Sub-G1 contents in HL-60 cells by NOMH90
Anti-cancer effect of NOMH90 evaluated by measuring Sub-G1 DNA content population percentage of the HL-60 cells after incubated with different concentrations of NOMH90 for 24-h. According to the results, NOMH90 fraction showed cell cycle arrest and accumulation of cells in the Sub-G1 phase in a dose-dependent manner (Fig. 6). Moreover, the apoptosis body formations corresponded with the accumulation of Sub-G1 content and further were determined as 33.2, 56.7, and 61.2% of Sub-G1 population against the NOMH90 concentrations (6.25, 12.5, and 25 μg mL−1), compared to the control, respectively (Fig. 6). Thus, we can suggest that NOMH90 has potential to induce cells death in HL-60 via inducing apoptosis in the nucleus.
DISCUSSION
Currently many studies focus on bio-diesel production and / or extract from the marine microalga N. oculata, (Converti et al. 2009, Umdu et al. 2009, Van Vooren et al. 2012) but limited interest on bioactivities of extracts from N. oculata microalga. The present study was undertaken to examine the anti-inflammatory effect of N. oculata extract and its sterol-rich fractions on LPS-stimulated RAW 264-7 cells and the anticancer effect on five cancer cell lines. We found that NOMH90 dose-dependently reduced the NO production in the cells and showed anticancer effect on HL-60 cancer cell line (Fig. 4). Therefore, we measured apoptosis body formation and cell cycle analysis only for NOMH90 using HL-60 cells. According to the apoptosis results and cell cycle analysis it clearly demonstrated that the reduction of cell viability of HL-60 cells are due to the cell death via induction of apoptosis pathways. Instead of above observations we found that anti-inflammatory and anti-cancer activities are quite similar to the sterol contents of the NOMH fractions.
Macrophage cells are considered as the first line of host defense against inflammation. When macrophage cells expose to the stimulating agents such as LPS, they start to express inflammatory responses to act against inflammatory stimuli (Smith 1994, Kim et al. 2006, Ma and Quirion 2008). iNOS and COX-2 are known as respective inflammatory mediators involved in the pathogenesis of diseases associated with inflammation (Lee et al. 2003, Kim et al. 2009). Expression of iNOS proteins induce the NO production and COX-2 induce the PGE2 production in LPS-activated macrophages cells (Han et al. 2004, Greenhough et al. 2009). Moreover, NO is a labile gaseous free radical as well as NO can act as a neurotransmitter, vasodilator and in cellular level defense against tumor cells, parasites, and bacteria in the immunological system (Nakagawa and Yokozawa 2002). Besides excess and prolong production of NO cause pathogenesis of inflammation, and cancer (Kobuchi et al. 1997). PGE2 is also capable to regulate platelet aggregation, vascular permeability, and thrombus formation in the development of inflammatory responses (Knöferl et al. 2001). Therefore, it is important to control the expression of iNOS and COX-2 to control inflammatory responses. Recently, number of studies confirmed that the compounds and solvent fractions separated from microalgae have potential to use as anti-inflammatory agents (Kim et al. 2013, Somchit et al. 2014). Instead of that, Yasukawa et al. (1996) reported that the sterols isolated from Chlorella vulgaris have promising activity against 2-O-tetradecanoylphorbol-13-acetate (TPA)-induced inflammation in mice (Yasukawa et al. 1996). Our present results also, indicated that NOMH90 (sterol-rich fraction) inhibited the NO production in LPS-stimulated RAW 264.7 cells. The anti-inflammatory effects of NOMH90 were accomplished by the decrease in expression of iNOS and COX-2 proteins in cytosol dose-dependent manner. Thus, there will be a potential to develop anti-inflammatory drugs using N. oculata microalga.
Keeping a balance between cell proliferation and cell death is an important mechanism to maintain homeostasis in multicellular organisms (Vanden Berghe et al. 2013). Induction of apoptosis is one major form of cell death and it can be observed from embryonic development through immunological regulation. Also, apoptosis is a useful tool to eliminate superfluous, inappropriate or harmful cells from the infected site in case of pathological situation (Jacobson et al. 1997). Deregulated levels of apoptosis in many cancer cells is a common problem to eliminate cancer cells in an effective manner. Therefore, the compounds that are capable of restoring apoptosis is known to be a promising strategy for cancer prevention (Fesik 2005). Justo et al. (2001) reported anticancer effect of extract collected from microalga Chlorella vulgaris. Further, their results demonstrated prolonged the survival of mice inoculated with the Ehrlich ascites tumor after administration of C. vulgaris extracts on Ehrlich ascites tumor infected mice compared to the control (Justo et al. 2001). According to our results (Figs 5 & 6) it is also indicated that NOMH90 fraction has great impact on inducing apoptosis in HL-60 cells in a dose dependent manner.
In conclusion, the present study highlighted the anti-inflammatory and anticancer activity of partially purified sterol-rich fraction isolated from 80% methanol extract and followed by open silica column chromatography of N. oculata microalga. According to the results of NOMH90 fractions of N. oculata microalga may be a feasible therapeutic or preventive candidate for developing drugs to treat diseases associated with inflammation and Human promyelocytic leukemia cancer.