INTRODUCTION
Gelidioid red algae are well-known raw materials yielding high-quality bacteriological and pharmaceutical-grade agar and agarose and having valorizing bioactivity (Rhein-Knudsen et al. 2015, Matos et al. 2020). Biocellulose nanocomposites can be developed from the microfibrillar material in the rhizines of Gelidium elegans (Chen et al. 2016). However, the recent shortage of gelidioid biomass due to overharvesting in nature is a major challenge to the future of the high-quality agar and agarose industries (Santos and Melo 2018). Bacterial agarose medium from gelidioids cannot be substituted by carrageenan or other products from red algae (Callaway 2015). There has been recognition of the need for sustainable supplies and investigations of biomass (Akatsuka 1986, Felicini and Perrone 1994, Friedlander 2008, Santos and Melo 2018), but the mass cultivation methods for gelidioids are still not developed.
Pterocladiella capillacea (S. G. Gmelin) Santelices & Hommersand, long referred to as Pterocladia capillacea (S. G. Gmelin) Bornet, is distributed globally from the Galápagos Islands on the equator to England in the northern hemisphere and New Zealand in the southern hemisphere (Freshwater et al. 1995, Boo et al. 2016). The plants grow by division of a dome-shaped apical cell, and they reach up to 30 cm in nature (Felicini and Perrone 1994). Although P. capillacea has a Polysiphonia-type life history, asexual tetrasporophytes predominate sexual carposporophytes in the field (Bottalico et al. 2008, Patarra et al. 2020). For example, in Baja California, Mexico, gametophytes occurred solely in January with a very low percentage (less than 0.15%) (Servièrse-Zaragoza and Scrosati 2000) and they have been reported to be rare in temperate waters (Felicini and Perrone 1994). Erect plants are mostly annual, while prostrate stolons are perennial, existing for 2–3 years (Felicini and Perrone 1994, Stewart 1968).
P. capillacea may be a suitable species for vegetative propagation due to its high capacity for regeneration from wounded branches (Felicini and Perrone 1994). Many researchers have studied the growth of P. capillacea under various conditions of temperature and irradiance in laboratories, providing valuable data about increasing biomass (Nasr et al. 1966, Friedlander and Zelikovitch 1984, Stewart 1984, Fralick et al. 1990, Macler and Zupan 1991, Yokoya and Oliveira 1992, Felicini et al. 2002, Gal-Or and Israel 2004, Harb et al. 2018, Patarra et al. 2019). However, their studies have focused mostly on laboratory cultivation. The aquaculture of P. capillacea in the sea or outdoor tanks has not made progress due to the poor survival and growth of tetraspore germlings as well as slow growth (Stewart 1984, Friedlander 2008, Santos and Melo 2018). The aim of the present work was to produce vegetative seedstock for mass cultivation of P. capillacea. We examined the number and length of regenerated branches arising from both cut edges of sectioned fragments in laboratory culture. This study is the first endeavor to cultivate P. capillacea in Korea, one of the major countries harvesting the species for agar extraction.
MATERIALS AND METHODS
Samples for growth experiments were collected from March 12–14, 2021, from the intertidal rocky shore at Dongbaek-ri, Gijang on the southeastern coast of Korea. Collections were transported to the laboratory within 20 min and kept in a tank of 10 L set with running seawater. The plants were cleaned, removing epibionts and surface sediments by a brush and knife under a compound microscope. They were rinsed repeatedly under running seawater for 2 days.
Healthy plants were selected by viewing under a compound microscope, and then clean and young branches were cut to 0.8 cm long using a medical disposable biopsy punch (Kai Industries Co., Ltd., Saitama, Japan). Growth experiments were carried in a programmable multi-room incubator (JSR, JSMI-04CP; JS Research Inc., Gongju, Korea) with light intensity set at about 27 μmol photons m−2 s−1 and a photoperiod of 10 dark: 14 light. Four temperature conditions were used; 10, 15, 20, and 25°C, which are approximately similar to seasonal seawater temperatures on Korean coasts (http://iridl.ldeo.columbia.edu/maproom/Global/Ocean-Temp/Monthly_Temp.html). Cultures were kept in a six-well TC-treated multi-well cell culture plate (353046; Falcon, Miami, FL, USA) (Fig. 1). Each well had a single fragment, and a total of 42 sectioned fragments were kept in seven replicates at each of the four temperature options. Provasoli’s enriched seawater medium was used (Starr and Zeikus 1993) and the medium was renewed every week. The culture experiments lasted for 3 weeks, and then all cultured fragments were photographed using a Tech Xcam III camera (Techsan Co., Ltd., Seoul, Korea) attached to a compound microscope (SZX10; Olympus, Tokyo, Japan). The length of the longest branch from both cut edges of cultured fragments was measured under the microscope. In addition, the number of branches from both cut sides was counted under the microscope.
Data from 40 fragments were used for statistical analysis except a couple of fragments with abnormal growth. However, 29 fragments were available at 10°C because of no growth of other fragments. All data were presented as mean ± standard error. We assessed the significance of the differences in the length of the longest regenerated branches and the number of regenerated branches on both sides of sectioned fragments among the different temperatures using ANOVAs. The analyses were based on a one-way design among four levels of temperatures as fixed factor. The means were compared using the F-test to determine which of the cultures differed significantly. When an ANOVA identified a significant difference (p < 0.05), Tukey’s post hoc multiple comparisons were used to identify where the differences occurred. All data analyses were performed using a jamovi 1.8.2 (http://www.jamovi.org).
RESULTS
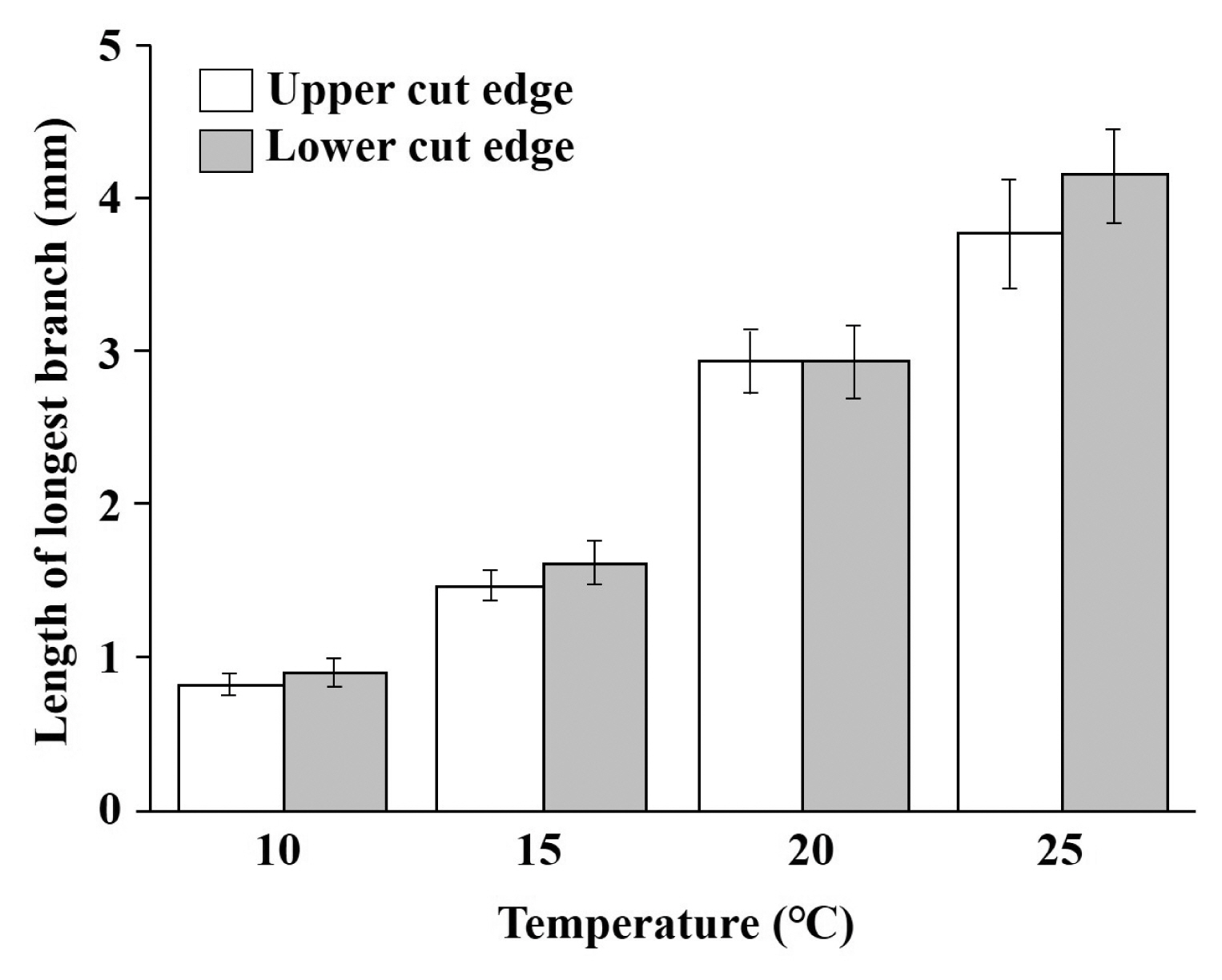
In all temperature conditions, sectioned fragments formed axis-like branches from the upper cut edge and stolon-like branches from the lower cut edge. Axis-like branches were mostly compressed and often had blunt tips (Fig. 2A–L). The length of axis-like branches averaged 0.84 ± 0.05 mm at 10°C cultivation and 1.47 ± 0.09 mm at 15°C. They became branched and long at 20°C (Fig. 2E & F) and the length increased substantially to 2.93 ± 1.28 mm. Their size averaged 3.77 ± 0.35 mm at 25°C. Stolon-like branches were mostly terete and often had acute tips (Fig. 2F). The length of solon-like branches averaged 0.92 ± 0.11 mm at 10°C and 1.63 ± 0.13 mm at 15°C. The length increased substantially to 2.93 ± 0.20 mm at 20°C and 4.15 ± 0.31 mm at 25°C (Fig. 3).
The number of axis-like branches averaged 2.13 ± 0.24 at 10°C and 2.58 ± 0.21 at 15°C. It increased to 2.74 ± 0.20 at 20°C but decreased to 2.56 ± 0.21 at 25°C (Fig. 4). The number of stolon-like branches averaged 1.42 ± 0.18 at 10°C. It increased substantially to 4.18 ± 0.44 at 15°C and 4.26 ± 0.44 at 20°C. However, it decreased to 2.64 ± 0.22 at 25°C (Fig. 4).
ANOVAs showed significant differences among cultures of sectioned fragments at all temperatures except the number of regenerated branches from the upper cut edge (Table 1). Temperature responses to all growth responses, represented as the length and number of regenerated branches, were significant (p < 0.001). However, the number of regenerated branches from the upper edge was not significant. In addition, growth responses to all temperature options were also significant (p < 0.001).
DISCUSSION
Our study reveals that the upper cut edge of sectioned fragments extends the regeneration area, favoring more axis-like branches rather than a single branch from a single apical cell of the Gelidiales. The lower cut edge also produces more stolon-like branches which helps in producing abundant rhizoids for anchoring to the substrata. The results reveal the high regeneration capacity and intrinsic bipolarity of P. capillacea, which is supported by the significant statistical differences in regeneration except the number of regenerated branches from the upper cut edge. The polarity from the sectioned fragments has been reported in previous studies on P. capillacea (Felicini and Perrone 1994, Felicini et al. 2002). Although the growth of fragments has been reported in many previous studies (Stewart 1984, Felicini and Perrone 1994, Gal-Or and Israel 2004), our study is the first on P. capillacea examining the number and length of regenerating branches from both the upper and lower cut edges of sectioned fragments. Axis-like regenerated branches may produce more biomass, while stolon-like branches may have more opportunity to produce holdfasts for anchoring to the substrate. Such fragments likely have a high chance of survival in the sea compared to spores or fragments lacking regenerated branches. A steady supply of fragments with regenerating branches may enable the bottleneck for cultivation of P. capillacea in the sea to be overcome.
Our experiments reveal that regeneration of Korean plants was good at 15–25°C. This is similar to Azorean plants in which growth and branch proliferation occurred at 20–22°C (Patarra et al. 2019). Yokoya and Oliveira (1992) reported that Brazilian plants were adapted to either 18°C or 22°C. Felicini et al. (2002) reported that the threshold temperature of distal shoots in fragment cultures of Italian plants was 19–24°C. However, the optimal growth temperature of P. capillacea likely depends on its geographical distribution.
P. capillacea is generally known as a sciophilous species (Gal-Or and Israel 2004, Harb et al. 2018). In our study, relatively low irradiance (27 μmol photons m−2 s−1) induced many regenerating branches at the optimal temperature range. Felicini et al (2002) reported that at 19–24°C, 20–40 μmol photons m−2 s−1 induced a compressed axis of Italian plants while 2.5–10 μmol photons m−2 s−1 induced a terete axis, similar to prostrate branches. High irradiances can improve the growth rates of P. capillacea, for example at 177 μmol photons m−2 s−1 for Azorean plants (Fralick et al. 1990) and 300 μmol photons m−2 s−1 for Brazilian plants (Harb et al. 2018). However, high irradiances may not favor mass cultivation because they boost the growth of epibionts during culture.
Similar propagation by fragmentation was reported in Gelidium (Fei and Huang 1991, Rodríguez 1996, Nelson and Farr 2003). Fei and Huang (1991) described regeneration of fragments (with section sizes of 2–5 mm) of G. elegans Kützing from China (as G. amansii), but it took 30 days to produce regeneration buds of 0.5–1 mm and 75 days for them to develop into creeping branches of 7 mm. We also found that the growth of new branches from sectioned fragments of G. elegans was very slow (unpublished data). Rodríguez (1996) reported vegetative propagation by fragmentation of G. gonzalezii Rodríguez et al. from Mexico (as G. sclerophyllum), in which growth in length and branch production were favored from medial fragments and rhizoid formation was favored from apical fragments. However, G. crinale subsp. longipes (J. Agardh) W. A. Nelson & G. H. Boo from New Zealand (as G. longipes) may be a suitable candidate for fragment propagation due to its capacity to regenerate from cut surfaces as well as producing abundant rhizoids for anchoring to the substrate (Nelson and Farr 2003). Vegetative reproduction through fragmentation is not uncommon in nature for red algae such as Dipterocladia, Gigartina, and Hypnea (Buggeln 1981, Cecere et al. 2011, Hwang and Boo 2019).
Our study reveals that 3 weeks’ cultivation of sectioned fragments at 15–25°C is a time-saving and feasible strategy to produce vegetative seedstock for mass cultivation. Fragments bearing regenerated branches can be transferred to outdoor tanks. Further research is needed to evaluate their transfer to rocky shore habitats, as well as their survival rate both in tanks and in situ compared to that of spores released in nature.
In addition to need for sustainable biomass for agar industries, there is considerable interest globally in the issue of coastal restoration and ocean afforestration, including the roles of subcanopy taxa (Tait and Schiel 2018). The development of appropriate seedstock and nursey techniques are essential for progress to meet these targets. We are continuing research on the use of sectioned fragments with regenerated branches for cultivation in ourdoor tanks or in the sea. Further methodological improvements, using sectioned fragments as seedstock, have potential to support agar industries in the near future, and may be able to be applied to other Gelidiales species that are low in regeneration capacity but high in agar quality.