Shin, Xing, Park, Yarish, Kong, and Kim: Effects of biostimulants, AMPEP and Kelpak on the growth and asexual reproduction of Pyropia yezoensis (Bangiales, Rhodophyta) at different temperatures
ABSTRACT
Acadian marine plant extract powder (AMPEP) and Kelpak are commercial biostimulants derived from brown algae Ascophyllum nodosum. This study was to determine if AMPEP and Kelpak can induce thermal resistance in Pyropia yezoensis. P. yezoensis blades were exposed to different concentrations (control: 0, low: 0.001, high: 1 ppm) of AMPEP and Kelpak at 10°C for 6 and 7 days, respectively. Those blades were then cultivated in von Stosch enriched seawater medium at different temperatures (10, 15, 20, and 25°C) with 12 : 12 L : D photoperiod and 100 μmol m−2 s−1 of photosynthetically active radiation for additional 15 days. Results showed that P. yezoensis reproduced archeospores at 20 and 25°C at all biostimulant conditions within 15 days. At lower temperatures (10 and 15°C), only AMPEP-treated P. yezoensis reproduced archeospores. P. yezoensis exposed to 1 ppm Kelpak exhibited higher phycoerythrin and phycocyanin contents than control and 0.001 ppm conditions at 15°C. AMPEP-treated conditions showed higher phycoerythrin and phycocyanin contents than control at 10°C. These results suggest that AMPEP and Kelpak may not enhance the thermal resistance of P. yezoensis. However, AMPEP stimulated archeospores release at lower temperatures. The treatment of AMPEP and Kelpak also increased the pigment contents in P. yezoensis. These results suggest that the use of seaweed-derived biostimulants can provide some economic benefits in P. yezoensis aquaculture. The enhancement of archeospores formation by AMPEP at lower temperature may also increase the productivity since Pyropia farming relies on the accumulation of secondary seedings via asexual reproduction.
Key words: AMPEP; Archeospores; asexual reproduction; biostimulant; Kelpak; Pyropia; temperature
INTRODUCTION
Seaweed-derived biostimulants are mainly used in agriculture for an advance of the development of root capacities to improve nutrient use efficiency ( Rayorath et al. 2008), to enhance resistance to abiotic stresses (e.g., frost and salinity) and to decrease the impacts of biotic stresses (e.g., pests, fungal diseases, and insect infestation) ( Jayaraj et al. 2008, Rayorath et al. 2008). One of the most widely known seaweed-based biostimulants is Kelpak (Kelp Products Pty Ltd., Simon’s Town, South Africa), which is a liquid biostimulant derived from the brown alga, Ecklonia maxima (Osbeck) Papenfuss ( Van Staden et al. 1995). Kelpak is known to improve the growth rate by stimulating phytohormones contents of plants ( Featonby-Smith and Van Staden 1983, 1987). For example, Kelpak can improve yield of strawberry ( Masny et al. 2004), fruit number, and size of pepper ( Arthur et al. 2003). Higher plants, Pelargonium peltatum and Brassica oleracea treated with Kelpak showed significantly higher chlorophyll (Chl) a and Chl b contents than the treatment without this biostimulant ( Krajnc et al. 2012, Rengasamy et al. 2016). Another higher plant species, Festulolium braunii, exposed to Kelpak also showed higher chlorophyll contents than control and therefore increased leaf greenness ( Sosnowski et al. 2019). To seaweed, Kelpak increased the growth of Gracilaria and Ulva ( Robertson-Andersson et al. 2006) and also enhanced thermal tolerance of Saccharina ( Umanzor et al. 2020b). Another commercial seaweed-derived biostimulant, Acadian marine plant extract powder (AMPEP; Acadian Seaplants Ltd., Dartmouth, Nova Scotia, Canada) is made from the brown alga, Ascophyllum nodosum (Linnaeus). Ascophyllum nodosum extract is known to improve yield and nutritional quality of spinach ( Fan et al. 2013) and yield and fruit quality of grapes ( Norrie et al. 2002). Also, it improves phenolics, antioxidant, and chlorophyll contents ( Norrie et al. 2002). In case of seaweed, AMPEP reduces epiphyte loads, and enhances growth and thermal tolerance on Kappaphycus ( Hurtado and Critchley 2018). Also, AMPEP treatment increased the biosynthesis of carrageenan in Kappaphycus (i.e., yield, viscosity, and gel strength) ( Ali et al. 2018). The growth of Gracilaria corticata and G. salicornia was also enhanced by AMPEP ( Dawange and Jaiswar 2020, Jaiswar et al. 2021).
Pyropia cultivation usually utilizes zygotospores from fertilized thalli to produce conchocelis, which grows in oyster shells ( Kim et al. 2022). This process demands space, time, and cost, and may also lead to diseases such as yellow spot and white spot ( Ryu et al. 2001, Guan et al. 2013). Germination and growth of archeospores of P. yezoensis are faster than conchospores ( Li 1989). Archeospores release amplifies crop biomass compared with conchospores seeding ( Li and Cui 1980, Wang 1985). Archeospores require only 1 or 2 days to develop into the two-cell stage, whereas protoplasts from blade need 5 to 7 days. Also, growth of gametophytes via archeospores enables the rapid multiplication of individuals ( Waaland et al. 1990, Chen et al. 1994). Archeospores rapidly attach to seeding ropes and develop into secondary stage ( Duffield et al. 1972). This suggests that methods utilizing asexual reproduction may be more efficient in terms of time, labor, space, and cost. This study was to test the effect of Kelpak and AMPEP on P. yezoensis at different temperatures and to determine the optimal concentrations of Kelpak and AMPEP for thermal resistance.
MATERIALS AND METHODS
Algal materials
The strain of P. yezoensis (PY-JB-ST1) used in this study was originally collected from Mabawi, Jebu Island, Gyeonggi-do, Korea (37°15′69.07″ N, 126°61′53.80″ E) in February 2018. After the collection, P. yezoensis blades were cut into 0.5 cm and cultivated under 100 μmol m−2 s−1 at 20°C to stimulate archeospores release. The gametophytic blades from archeospores were then cultivated at 10°C, 12 : 12 L : D photoperiod and 80 μmol m−2 s−1 at the Marine Ecology and Green Aquaculture Laboratory, Incheon National University, Korea.
Experimental design
Kelpak and AMPEP experiments were conducted independently. The full profile of nutrients of these biostimulants can be found in Hurtado et al. (2009) and Lötze and Hoffman (2016). P. yezoensis blades (1 cm × 1 cm) were cultured at 0.1 g L −1 stocking density at 10°C in different concentrations (control: 0, low: 0.001, high: 1 ppm) of Kelpak and AMPEP for 7 and 6 days, respectively. Photosynthetically active radiation (PAR) and photoperiod were 90 ± 10 μmol m −2 s −1 and 12 : 12 L : D, respectively. Nutrients were supplied by von Stosch enriched (VSE) medium, and 250 mg L −1 of germanium dioxide (GeO 2) was added to inhibit the development of diatoms ( Lewin 1966). To prevent nutrient limitation, VSE medium was added after 4 days. After exposure to Kelpak or AMPEP, all the blades were rinsed with filtered and sterilized seawater and transferred to 100 mL flask with VSE seawater at 0.5 g L −1 stocking density, 30 psu salinity, 12 : 12 L : D photoperiod and 90 ± 10 μmol m −2 s −1 of PAR. P. yezoensis blades were then cultivated at four different temperatures, 10, 15, 20, and 25°C for additional 15 days. Continuous aeration was provided and the culture medium with VSE was changed every 5 days. To prevent nutrient limitation, VSE was added to the culture medium 3 days after the medium change.
Measurements of growth rate and pigments
Fresh weight (FW) of P. yezoensis in each cultivation condition was measured every 5 days. Water on the blades was removed with paper towels before weight measurement. Weight of each condition was restored to the initial stocking density after the measurement. The specific growth rate (SGR) was calculated by the following formula ( Kim et al. 2007): , where S1 and S2 are the initial and final FW on days T1 (initial day) and T2 (final day), respectively.
For phycoerythrin (PE) and phycocyanin (PC) analyses, approximately 0.05 g (FW) of fresh thalli was ground in 0.1 M phosphate buffer (pH 6.8). After centrifugation at 5,000 ×g for 15 min at 4°C, the absorption of supernatant was measured for PE and PC following Beer and Eshel (1985):
Statistical analysis
All statistical analyses were conducted using IBM SPSS Statistics version 27 (IBM, Armonk, NY, USA). Two-way ANOVA were performed to determine the effects of biostimulants concentrations and temperatures on SGR and PE and PC contents. Tukey’s honest significant difference tests were conducted for post-hoc investigation. Differences between treatments were considered to be significant at p < 0.05.
RESULTS
Effect of Kelpak on Pyropia yezoensis
SGRs of P. yezoensis were significantly influenced by Kelpak (p = 0.047), temperature (p < 0.001) and the interaction of these two factors (p = 0.002) ( Table 1). P. yezoensis grew well in a wide range of temperatures (10, 15, 20, and 25°C) for 10 days ( Fig. 1). There was no significant difference in the SGR at 10–20°C. At control without Kelpak, the SGR at 15°C was significantly higher than that at 25°C. However, at 1 ppm, the SGR was similar at all temperature conditions. The lowest growth rate was observed at 0.001 ppm and 25°C ( Table 2). Archeospores release was observed in all conditions regardless of the Kelpak concentrations at 20 and 25°C in 15 days ( Fig. 2). PE and PC contents of P. yezoensis were measured at 10 and 15°C only due to asexual reproduction at 20 and 25°C. PE content was significantly influenced by temperature (p = 0.008), Kelpak (p = 0.008), and the interaction of these two factors (p < 0.001) ( Table 1). No significant effects of Kelpak were observed in PE content at 10°C. At 15°C, however, PE content at 1 ppm was significantly higher than control and 0.001 ppm of Kelpak, and similar to other conditions at 10°C ( Fig. 3A). PC content was significantly affected by temperature (p < 0.001) and Kelpak (p = 0.010) ( Table 1). PC content showed a similar pattern to PE content. No significant differences were observed in PC content at 10°C while the content in 1 ppm was the highest at 15°C ( Table 3, Fig. 3B).
Effect of AMPEP on Pyropia yezoensis
SGR of P. yezoensis was significantly influenced by temperature (p < 0.001), whereas AMPEP and the interaction of temperature and AMPEP did not affect significantly (p > 0.05) ( Table 1). Regardless of the AMPEP concentrations, the lowest growth rate was observed at 25°C ( Fig. 4). The SGR at control and 0.001 ppm was significantly lower at 20 than 15°C. Archeospores release was observed at 20 and 25°C regardless of the AMPEP concentrations in 15 days. At 10 and 15°C, archeospores release was also observed in AMPEP-treated conditions ( Fig. 5). Like the Kelpak experiment, PE and PC contents of P. yezoensis were measured at 10 and 15°C. PE content was significantly influenced by temperature (p = 0.001), AMPEP (p = 0.002), and the interaction of these two factors (p = 0.001) ( Table 1). PE content in AMPEP-treated thalli was significantly higher than that in control at 10°C. There was no significant differences in PE content at 15°C ( Fig. 6A). PC content was significantly affected by temperature (p = 0.001), concentration of AMPEP (p = 0.010) and the interaction of these two factors (p = 0.009) ( Table 1). PC content showed a similar pattern to PE content. PE content in 0.001 ppm at 10°C was significantly higher than those in other conditions ( Table 3, Fig. 6B).
DISCUSSION
Asexual reproduction (production of archeospores) occurred in P. yezoensis at high temperature conditions (20 and 25°C) in 15 days at all AMPEP and Kelpak concentrations. Archeospores were also observed in P. yezoensis treated with AMPEP in 15 days, even at 10 and 15°C, but was not observed in the samples pre-treated with Kelpak. These results suggest that AMPEP may enhance asexual reproduction in P. yezoensis. The formation of archeospores from P. yezoensis is influenced by various factors such as temperature, light intensity, wound, blade size, and desiccation ( Li 1989, Notoya et al. 1993, Suda and Mikami 2020). In the present study, higher temperature (20 and 25°C) clearly stimulated the release of archeospores. The promotion of the archeospores at higher temperature (20–24°C) was previously observed in the red alga Bangia fuscopurpurea ( Wang et al. 2008). Pyropia lacerata and Porphyra suborbiculata also released archeospores at 20 and 25°C ( Notoya et al. 1993). However, P. yezoensis cut to about 1 mm 2 formed callus cells, which are a three-dimensional cluster of cells randomly divided, without the formation of archeospores ( Suda and Mikami 2020). The callus cells germinated into gametophytic thalli like those of archeospores ( Suda and Mikami 2020). The present study cut the blade into 1 cm 2 in area, and all blades formed archeospores at higher temperatures. This result suggests that the blade size is an important factor to form archeospores. At lower temperatures, 10 and 15°C, asexual reproduction was observed when treated with AMPEP, suggesting that AMPEP influences the asexual reproduction of P. yezoensis. In plants, the extract of A. nodosum induces higher levels of abscisic acid (ABA) and Ca 2+ contents ( Wally et al. 2013, Shukla et al. 2018). Previous studies have shown that Ca 2+ influx and exposure to H 2O 2 and allantoin can promote archeospores release in P. yezoensis ( Takahashi et al. 2010). H 2O 2 has shown to facilitate the influx of Ca 2+ ( Gui et al. 2022), while allantoin can increase the levels of ABA and a phytohormone ( Watanabe et al. 2014). Therefore, AMPEP may help induce the release of archeospores in P. yezoensis through a similar mechanism. Asexual reproduction can help maintain desirable traits in the population, as the offspring are genetically identical to parental plants. This is beneficial for commercial cultivation for crops, as it ensures that the quality and taste remain consistent ( Li 1989, Waaland et al. 1990, Chen et al. 1994). Asexual reproduction is the principal seeding for commercial Pyropia cultivation ( Wada 1941, Li 1984). Interestingly, new blades produced via asexual reproduction, when the parental plants were treated with AMPEP. These thalli showed higher growth rates than the control without AMPEP at 10°C, which is the optimal growth temperature for this species ( Umanzor et al. 2022b). These results suggest that AMPEP not only promotes asexual reproduction but also enhances the growth of new blades, which is advantageous for commercial aquaculture of Pyropia. This is the first study showing that the seaweed-derived biostimulants, Kelpak and AMPEP can enhance the PE and PC contents. PE and PC are pigment-protein complexes found in certain photosynthetic organisms, particularly in cyanobacteria (blue-green algae) and red algae. These complexes play a crucial role in light harvesting during photosynthesis ( Bryant 1982). Recent studies reported that Kelpak increased photosynthesis and chlorophyll fluorescence of plants ( Krajnc et al. 2012, Rengasamy et al. 2016). Umanzor et al. (2020b) observed a darker color on AMPEP-treated Saccharina sporophytes. Similar results were also found in Kappaphycus alvarezii thalli at 16°C ( Loureiro et al. 2014). The higher contents of PE and PC in P. yezoensis are indicative of the efficiency of photosynthesis. This is significant because photosynthesis is the primary process that drives growth and productivity in these seaweed ( Raven et al. 2014). The PE and PC have wide applications in food, cosmetics and pharmaceutical industries due to their natural optical and biological properties ( Kim et al. 2018). The pigment contents in P. yezoensis, specifically the amount per unit area or weight, is crucial in determining the quality due to a strong correlation between pigment content and the quality of dried P. yezoensis sheets ( Aruga 1984). Higher PE contents in Pyropia lead to higher free amino acids contents, which are known to be the most significant factors influencing the taste of Pyropia ( Yoshie et al. 1993). Therefore, exposure to AMPEP and Kelpak have the potential to enhance the market value of P. yezoensis by stimulating the synthesis of PE and PC.
Pyropia yezoensis exposed to Kelpak did not reproduce at 10 and 15°C. Whereas, AMPEP-treated samples released archeospores within 15 days following a 6-day pretreatment at 1 ppm. In contrast, Han et al. (2023) reported that P. yezoensis didn’t reproduce when exposed to the same condition for 10 days. In Eucheumatopsis isiformis, a longer exposure to AMPEP (28 days at 5 ppm) resulted in a positive effect compared to a shorter exposure (45 min) or no exposure. The longer exposure produced more lateral shoots in E. isiformis ( Umanzor et al. 2020a), indicating a pronounced stimulatory effect on lateral shoot formation by AMPEP. This result suggests that exposure time is a crucial factor determining the effects of biostimulants.
In conclusion, this study showed the potential of seaweed-derived biostimulants as a novel technology to improve P. yezoensis cultivation. AMPEP induced the formation and release of archeospores in P. yezoensis more efficiently than Kelpak, increasing production. Additionally, the use of Kelpak and AMPEP can increase PE and PC contents in this alga, resulting in higher quality products of P. yezoensis.
ACKNOWLEDGEMENTS
This research was supported by the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2017R1A6A1A06015181) and by the Ministry of Science and ICT (NRF-2022R1A2C1011394), and funded by the Ministry of Oceans and Fisheries of Korea (Project No. 20190518).
CONFLICTS OF INTEREST
The authors declare that they have no potential conflicts of interest.
Fig. 1
Specific growth rates (SGRs) of Pyropia yezoensis in different Kelpak concentrations and temperatures at day 15. Each coordinate is the overall mean ± standard deviation (STD) of 9 replicates measured at 10 and 15°C. Since reproduction occurred at 20 and 25°C within 15 days, 6 replications were used for the analysis at these temperatures. Different letters on the bars indicate significant differences (p < 0.05). Error bars represent mean ± STD of 6 or 9 replicates. 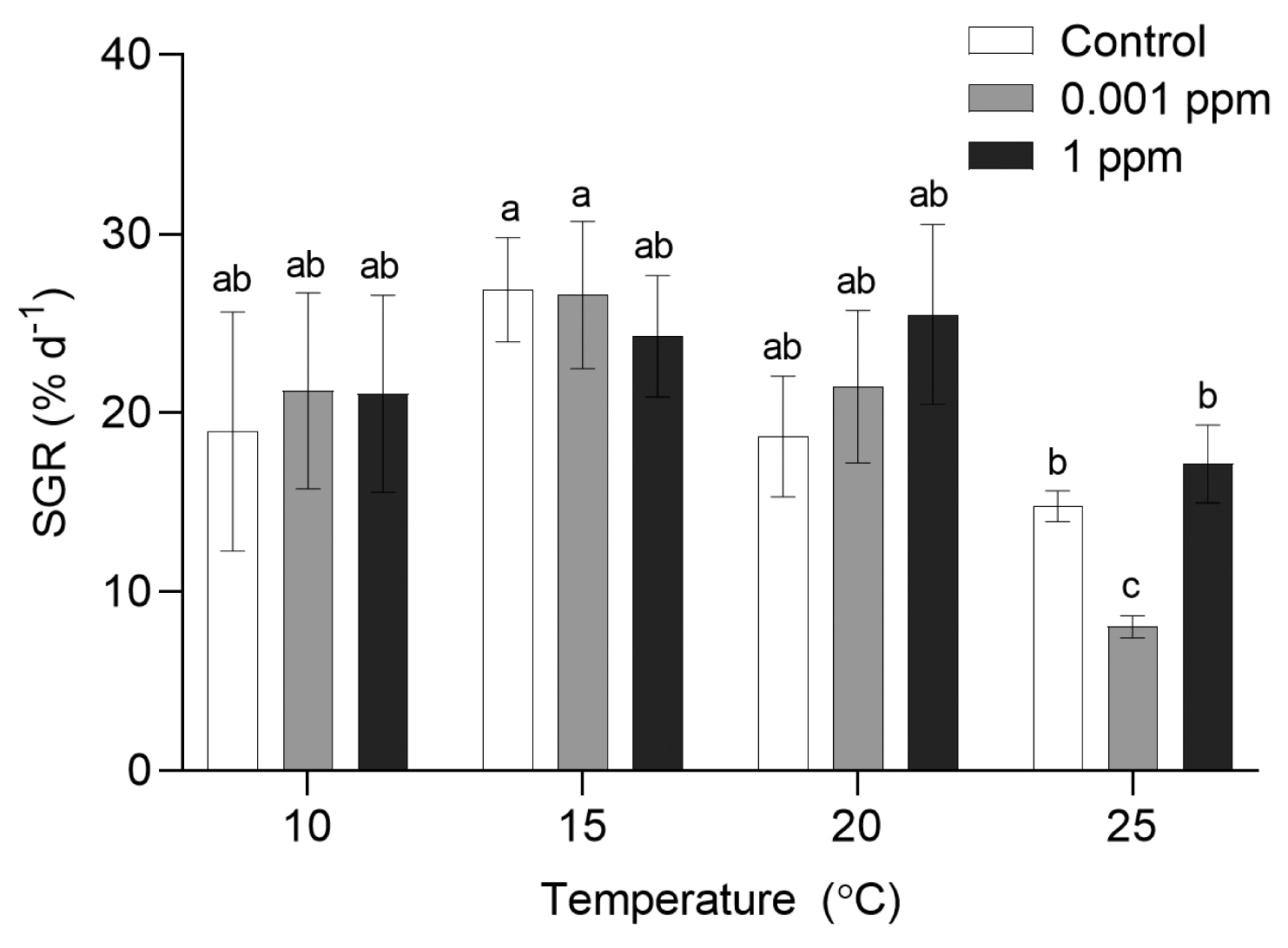
Fig. 2
Microscopic images of gametophytes of Pyropia yezoensis after 15 days of cultivation at 20 and 25°C at different Kelpak and temperatures. (A) Control at 20°C. (B) 0.001 ppm of Kelpak at 20°C. (C) 1 ppm of Kelpak at 20°C. (D) Control at 25°C. (E) 0.001 ppm of Kelpak at 25°C. (F) 1 ppm of Kelpak at 25°C. Scale bars represent: A–F, 100 μm. 
Fig. 3
Phycoerythrin (A) and phycocyanin (B) contents of Pyropia yezoensis at different temperatures (10 and 15°C) and different concentrations of Kelpak at day 15. FW, fresh water. Different letters on the bars indicate significant differences (p < 0.05). Error bars represent mean ± standard deviation of 9 replicates. 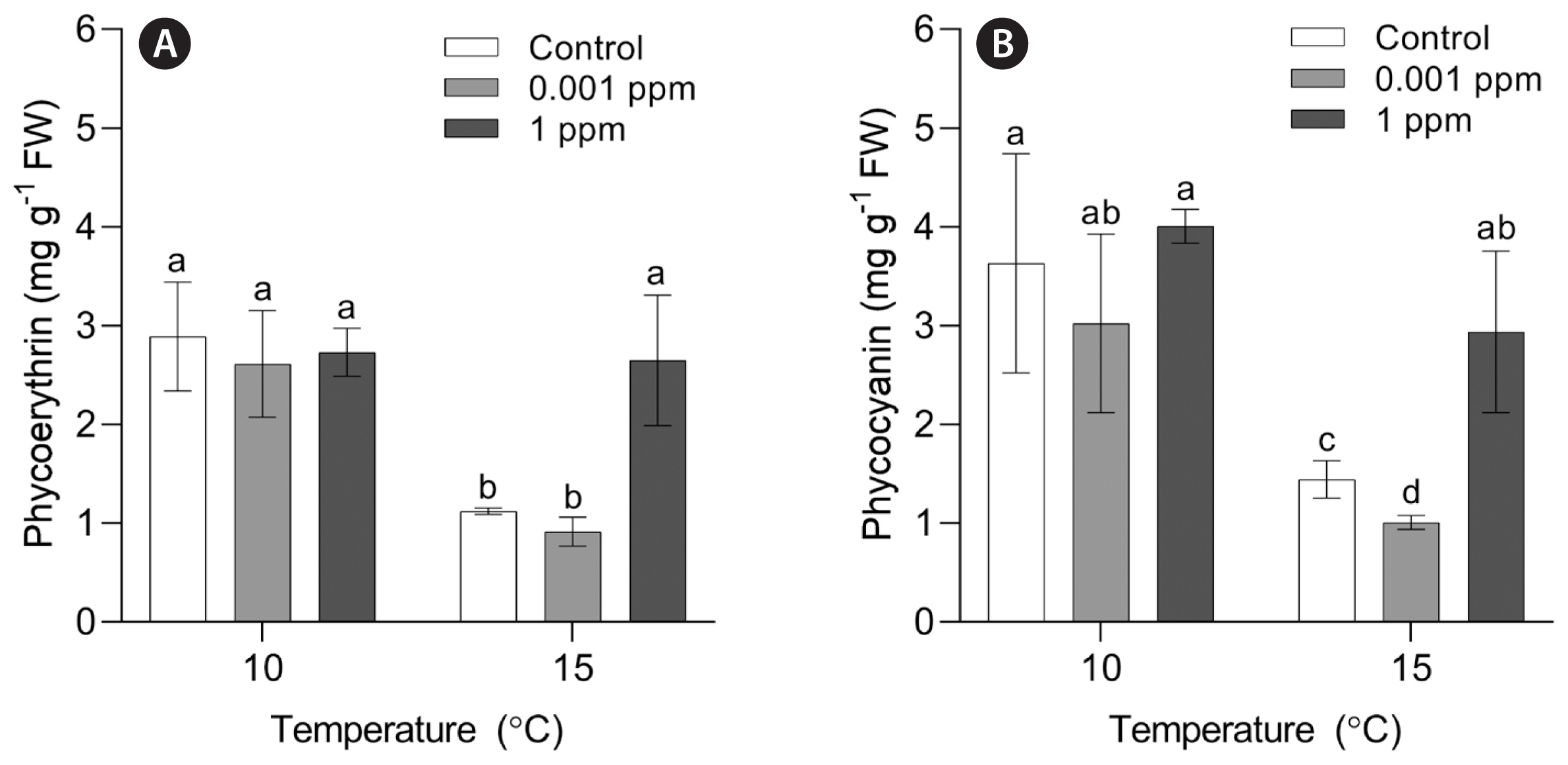
Fig. 4
Specific growth rates (SGRs) of Pyropia yezoensis in different Acadian marine plant extract powder (AMPEP) concentrations and temperatures at day 15. Each coordinate is the overall mean ± standard deviation (STD) of 9 replicates measured at 10 and 15°C. Since reproduction occurred within 15 days, 6 replications were used for the conditions at 20 and 25°C. Different letters on the bars indicate significant differences (p < 0.05). Error bars represent mean ± STD of 6 or 9 replicates. 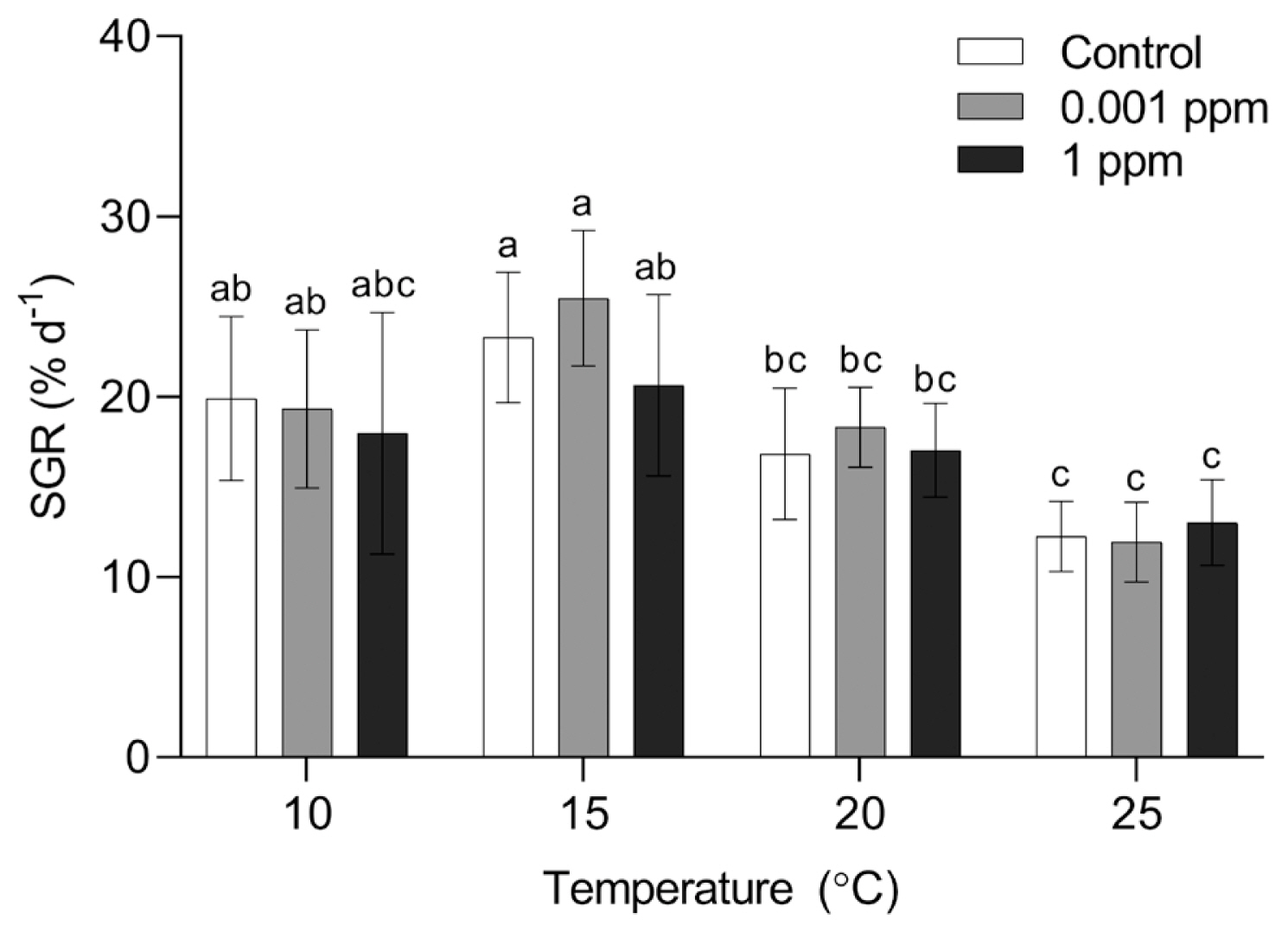
Fig. 5
Microscopic images of gametophytes of Pyropia yezoensis after 20 days (A–D) and 15 days (F, G, I & J) of cultivation at different Acadian marine plant extract powder (AMPEP) and temperatures. (A) 0.001 ppm of AMPEP at 10°C. (B) 1 ppm of AMPEP at 10°C. (C) 0.001 ppm of AMPEP at 15°C. (D) 1 ppm of AMPEP at 15°C. (E) Control at 20°C. (F) 0.001 ppm of AMPEP at 20°C. (G) 1 ppm of AMPEP at 20°C. (H) Control at 25°C. (I) 0.001 ppm of AMPEP at 25°C. (J) 1 ppm of AMPEP at 25°C. Scale bars represent: A–J, 100 μm. 
Fig. 6
Phycoerythrin (A) and phycocyanin (B) contents in Pyropia yezoensis at different Acadian marine plant extract powder concentrations and temperatures (10 and 15°C) at day 15. FW, fresh water. Different letters on the bars indicate significant differences (p < 0.05). Error bars represent mean ± standard deviation of 9 replicates. 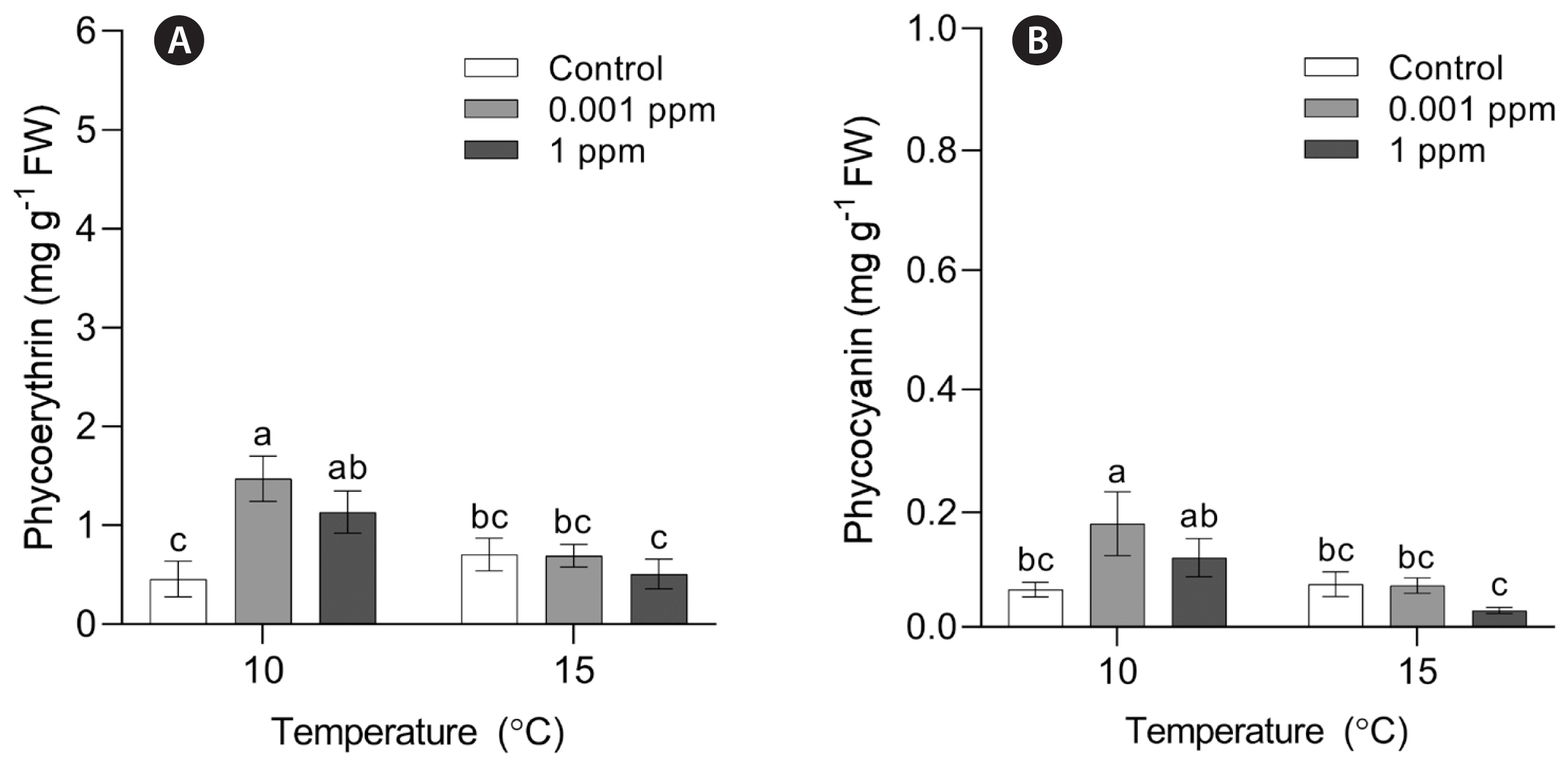
Table 1
Results of analysis of variance examining the effects of different biostimulants concentrations (0, 0.001, and 1 ppm) and different temperatures (10, 15, 20, and 25°C)
|
Source |
|
Concentration |
Temperature |
Concentration × temperature |
Error |
|
SGR (Kelpak) |
df |
2 |
3 |
6 |
60 |
|
F |
8.533 |
58.962 |
7.697 |
|
|
Sig. |
0.001
|
<0.001
|
<0.001
|
|
|
SGR (AMPEP) |
df |
2 |
3 |
6 |
60 |
|
F |
12.500 |
75.636 |
6.950 |
|
|
Sig. |
<0.001
|
<0.001
|
<0.001
|
|
|
Phycoerythrin (Kelpak) |
df |
2 |
1 |
2 |
12 |
|
F |
1.384 |
33.937 |
1.364 |
|
|
Sig. |
0.008
|
0.008
|
<0.001
|
|
|
Phycoerythrin (AMPEP) |
df |
2 |
1 |
2 |
12 |
|
F |
11.595 |
20.638 |
14.267 |
|
|
Sig. |
0.002
|
0.001
|
0.001
|
|
|
Phycocyanin (Kelpak) |
df |
2 |
1 |
2 |
12 |
|
F |
7.027 |
29.959 |
1.166 |
|
|
Sig. |
0.010
|
<0.001
|
0.345 |
|
|
Phycocyanin (AMPEP) |
df |
2 |
1 |
2 |
12 |
|
F |
6.914 |
21.613 |
7.268 |
|
|
Sig. |
0.010
|
0.001
|
0.009
|
|
Table 2
Specific growth rate of Pyropia yezoensis
|
Treatment |
Specific growth rate (% d−1) |
|
|
Control |
0.001 ppm |
1 ppm |
|
Kelpak |
|
|
|
|
10°C |
18.96 ± 6.67ab
|
21.24 ± 5.48ab
|
21.09 ± 5.51ab
|
|
15°C |
26.89 ± 2.90a
|
26.60 ± 4.11a
|
24.28 ± 3.39ab
|
|
20°C |
18.69 ± 3.38ab
|
21.46 ± 4.26ab
|
25.51 ± 5.02ab
|
|
25°C |
14.79 ± 0.85b
|
8.06 ± 0.63c
|
17.15 ± 2.17b
|
|
AMPEP |
|
|
|
|
10°C |
19.92 ± 4.54ab
|
19.23 ± 4.38ab
|
17.98 ± 6.70abc
|
|
15°C |
23.29 ± 3.63a
|
25.48 ± 3.76a
|
20.66 ± 5.02ab
|
|
20°C |
16.83 ± 3.63bc
|
18.32 ± 2.22bc
|
17.03 ± 2.60bc
|
|
25°C |
12.26 ± 1.95c
|
11.95 ± 2.22c
|
13.02 ± 2.38c
|
Table 3
Phycoerythrin and phycocyanin contents of Pyropia yezoensis
|
Phycoerythrin (mg g−1 FW) |
Phycocyanin (mg g−1 FW) |
|
|
|
Control |
0.001 ppm |
1 ppm |
Control |
0.001 ppm |
1 ppm |
|
Kelpak |
|
|
|
|
|
|
|
10°C |
2.89 ± 0.55a
|
2.61 ± 0.54a
|
2.73 ± 0.24a
|
3.63 ± 1.11a
|
3.02 ± 0.91ab
|
4.01 ± 0.17a
|
|
15°C |
1.12 ± 0.03b
|
0.91 ± 0.15b
|
2.65 ± 0.66a
|
1.44 ± 0.19c
|
1.01 ± 0.07d
|
2.93 ± 0.81ab
|
|
AMPEP |
|
|
|
|
|
|
|
10°C |
0.46 ± 0.18c
|
1.47 ± 0.23a
|
1.13 ±0.21ab
|
0.07 ± 0.01bc
|
0.18 ± 0.06a
|
0.12 ± 0.03ab
|
|
15°C |
0.71 ± 0.16bc
|
0.69 ± 0.11bc
|
0.51 ± 0.15c
|
0.07 ± 0.02bc
|
0.07 ± 0.01bc
|
0.03 ± 0.01c
|
REFERENCES
Ali, M. M., Sani, M. Z. B., Hi, K. K., Yasir, S. M., Critchley, A. T. & Hurtado, A. Q. 2018. The comparative efficiency of a brown algal-derived biostimulant extract (AMPEP), with and without supplemented PGRs: the induction of direct, axis shoots as applied to the propagation of vegetative seedlings for the successful mass cultivation of three commercial strains of Kappaphycus in Sabah, Malaysia. J. Appl. Phycol. 30:1913–1919.
doi.org/10.1007/s10811-017-1366-1
  Arthur, G. D., Stirk, W. A., van Staden, J. & Scott, P. 2003. Effect of a seaweed concentrate on the growth and yield of three varieties of Capsicum annuum. S. Afr. J. Bot. 69:207–211.
doi.org/10.1016/S0254-6299(15)30348-3
Aruga, Y. 1984.
In vivo absorption spectra and pigment contents of the two types of color mutants of Porphyra. Jpn. J. Phycol. 32:243–250.
Beer, S. & Eshel, A. 1985. Determining phycoerythrin and phycocyanin concentrations in aqueous crude extracts of red algae. Aust. J. Mar. Freshw. Res. 36:785–792.
doi.org/10.1071/MF9850785
Chen, LC-M, Craigie, J. S. & Xie, Z. K. 1994. Protoplast production from Porphyra linearis using a simplified agarase procedure capable of commercial application. J. Appl. Phycol. 6:35–39.
doi.org/10.1007/BF02185902
Dawange, P. & Jaiswar, S. 2020. Effects of Ascophyllum marine plant extract powder (AMPEP) on tissue growth, proximate, phenolic contents, and free radical scavenging activities in endemic red seaweed Gracilaria corticata var. cylindrica from India. J. Appl. Phycol. 32:4127–4135.
doi.org/10.1007/s10811-020-02254-6
Duffield, E. C., Waaland, S. D. & Cleland, R. 1972. Morphogenesis in the red alga, Griffithsia pacifica: regeneration from single cells. Planta. 105:185–195.
doi.org/10.1007/BF00385390
 Fan, D., Hodges, D. M., Critchley, A. T. & Prithiviraj, B. 2013. A commercial extract of brown macroalga ( Ascophyllum nodosum) affects yield and the nutritional quality of spinach in vitro. Commun. Soil Sci. Plant Anal. 44:1873–1884.
doi.org/10.1080/00103624.2013.790404
Featonby-Smith, B. C. & Van Staden, J. 1983. The effect of seaweed concentrate on the growth of tomato plants in nematode-infested soil. Sci. Hortic. 20:137–146.
doi.org/10.1016/0304-4238(83)90134-6
Guan, X., Li, J., Zhang, Z. & et al 2013. Characterizing the microbial culprit of white spot disease of the conchocelis stage of Porphyra yezoensis (Bangiales, Rhodophyta). J. Appl. Phycol. 25:1341–1348.
doi.org/10.1007/s10811-013-9976-8
Gui, T.-Y., Gao, D.-H., Ding, H.-C. & Yan, X.-H. 2022. Identification of respiratory burst oxidase homolog (Rboh) family genes from Pyropia yezoensis and their correlation with archeospore release. Front. Plant Sci. 13:929299
doi.org/10.3389/fpls.2022.929299
 Han, S., Song, H. I., Park, J. S. & et al 2023.
Sargassum horneri and Ascophyllum nodosum extracts enhance thermal tolerance and antioxidant activity of Neopyropia yezoensis. J. Appl. Phycol. 35:201–207.
doi.org/10.1007/s10811-022-02870-4
Hurtado, A. Q. & Critchley, A. T. 2018. A review of multiple biostimulant and bioeffector benefits of AMPEP, an extract of the brown alga Ascophyllum nodosum, as applied to the enhanced cultivation and micropropagation of the commercially important red algal carrageenophyte Kappaphycus alvarezii and its selected cultivars. J. Appl. Phycol. 30:2859–2873.
doi.org/10.1007/s10811-018-1407-4
Hurtado, A. Q., Yunque, D. A., Tibubos, K. & Critchley, A. T. 2009. Use of Acadian marine plant extract powder from Ascophyllum nodosum in tissue culture of Kappaphycus varieties. J. Appl. Phycol. 21:633–639.
doi.org/10.1007/s10811-008-9395-4
Jaiswar, S., Dawange, P. S., Thanth, C. & Mantri, V. A. 2021. Apical, sub-apical, and basal explants of industrially exploited marine red alga Gracilaria salicornia exhibited differential response to commercial seaweed-derived plant bio-stimulants. J. Appl. Phycol. 33:3975–3985.
doi.org/10.1007/s10811-021-02594-x
Kim, E.-Y., Choi, Y. H. & Nam, T.-J. 2018. Identification and antioxidant activity of synthetic peptides from phycobiliproteins of Pyropia yezoensis. Int. J. Mol. Med. 42:789–798.
doi.org/10.3892/ijmm.2018.3650
Kim, H.-S., Choi, H. G., Hwang, M.-S., Jeon, Y. J., Yarish, C. & Kim, J. K. 2022. Concise review of the genus Neopyropia (Rhodophyta: Bangiales). J. Appl. Phycol. 34:1805–1824.
doi.org/10.1007/s10811-022-02776-1
Kim, J. K., Kraemer, G. P., Neefus, C. D., Chung, I. K. & Yarish, C. 2007. Effects of temperature and ammonium on growth, pigment production and nitrogen uptake by four species of Porphyra (Bangiales, Rhodophyta) native to the New England coast. J. Appl. Phycol. 19:431–440.
doi.org/10.1007/s10811-006-9150-7
Kim, J. K., Yarish, C., Hwang, E. K., Park, M. & Kim, Y. 2017. Seaweed aquaculture: cultivation technologies, challenges and its ecosystem services. Algae. 32:1–13.
doi.org/10.4490/algae.2017.32.3.3
Krajnc, A. U., Ivanuš, A., Kristl, J. & Šušek, A. 2012. Seaweed extract elicits the metabolic responses in leaves and enhances growth of Pelargonium cuttings. Eur. J. Hortic. Sci. 77:170–181.
Li, S. 1989. Physio-ecological characteristics of monospores and monosporelings of Porphyra yezoensis Ueda and their application in cultivation. Stud. Mar. Sin. 30:81–92.
Li, S. & Cui, G. 1980. An observation on the growth and development of sporelings from conchospores and monospores of Porphyra yezoensis. Oceanol. Limnol. Sin. 4:370–374.
Li, S. Y. 1984. The ecological characteristics of monospores of Porphyra yezoensis Ueda and their use in cultivation. Hydrobiologia. 116/117:255–258.
doi.org/10.1007/BF00027679
Lötze, E. & Hoffman, E. W. 2016. Nutrient composition and content of various biological active compounds of three South African-based commercial seaweed biostimulants. J. Appl. Phycol. 28:1379–1386.
doi.org/10.1007/s10811-015-0644-z
Loureiro, R. R., Reis, R. P. & Marroig, R. G. 2014. Effect of the commercial extract of the brown alga Ascophyllum nodosum Mont. On Kappaphycus alvarezii (Doty) Doty ex P.C. Silva in situ submitted to lethal temperatures. J. Appl. Phycol. 26:629–634.
doi.org/10.1007/s10811-013-0085-5
Masny, A., Basak, A. & Żurawicz, E. 2004. Effect of foliar applications of Kelpak SL and Goëmar BM 86 preparations on yield and fruit quality in two strawberry cultivars. J. Fruit Ornam. Plant Res. 12:23–27.
Notoya, M., Kikuchi, N., Matsuo, M., Aruga, Y. & Miura, A. 1993. Culture studies of four species of Porphyra (Rhodophyta) from Japan. Nippon Suisan Gakkaishi. 59:431–436.
doi.org/10.2331/suisan.59.431
Rayorath, P., Jithesh, M. N., Farid, A. & et al 2008. Rapid bioassays to evaluate the plant growth promoting activity of Ascophyllum nodosum (L.) Le Jol. using a model plant, Arabidopsis thaliana (L.) Heynh. J. Appl. Phycol. 20:423–429.
doi.org/10.1007/s10811-007-9280-6
Rengasamy, K. R. R., Kulkarni, M. G., Pendota, S. C. & Van Staden, J. 2016. Enhancing growth, phytochemical constituents and aphid resistance capacity in cabbage with foliar application of eckol: a biologically active phenolic molecule from brown seaweed. N. Biotechnol. 33:273–279.
doi.org/10.1016/j.nbt.2015.11.002
Robertson-Andersson, D., Leitao, D., Bolton, J. J., Anderson, R. J., Njobeni, A. & Ruck, K. 2006. Can kelp extract (KELPAK) be useful in seaweed mariculture? J. Appl. Phycol. 18:315–321.
doi.org/10.1007/s10811-006-9030-1
Ryu, S., Cho, S., Park, S. & et al 2001. Cloning of the cel9A gene and characterization of its gene product from marine bacterium Pseudomonas sp. SK38. Appl. Microbiol. Biotechnol. 57:138–145.
doi.org/10.1007/s002530100743
Shukla, P. S., Shotton, K., Norman, E., Neily, W., Critchley, A. T. & Prithiviraj, B. 2018. Seaweed extract improve drought tolerance of soybean by regulating stress-response genes. AoB Plants. 10:plx051
doi.org/10.1093/aobpla/plx051
Sosnowski, J., Jankowski, K., Truba, M. & Malinowska, E. 2019. Effect of Ecklonia maxima extract on photosynthesis activity and chlorophyll content of Medicago × varia Martyn leaves. Chil. J. Agric. Res. 79:257–265.
doi.org/10.4067/S0718-58392019000200257
Suda, M. & Mikami, K. 2020. Reproductive responses to wounding and heat stress in gametophytic thalli of the red alga Pyropia yezoensis. Front. Mar. Sci. 7:394
doi.org/10.3389/fmars.2020.00394
Takahashi, M., Saga, N. & Mikami, K. 2010. Photosynthesis-dependent extracellular Ca 2+ influx triggers an asexual reproductive cycle in the marine red macroalga Porphyra yezoensis. Am. J. Plant. Sci. 1:1–11.
doi.org/10.4236/ajps.2010.11001
Umanzor, S., Han, S., Song, H.-I. & et al 2022a. Ascertaining the interactions of brown seaweed-derived biostimulants and seawater temperature on spore release, germination, conchocelis, and newly formed blades of the commercially important red alga Neopyropia yezoensis? Algal Res. 64:102692
doi.org/10.1016/j.algal.2022.102692
Umanzor, S., Han, S., Song, H.-I. & et al 2022b. Enhancements provided by the use of an Ascophyllum nodosum extract can be transferred through archeospores in the red alga Neopyropia yezoensis (Ueda). In : Yang L.-E., Brodie J., editors Aquat. Bot. 177:103481
doi.org/10.1016/j.aquabot.2021.103481
Umanzor, S., Jang, S., Antosca, R., Critchley, A. T., Yarish, C. & Kim, J. K. 2020a. Optimizing the application of selected biostimulants to enhance the growth of Eucheumatopsis isiformis, a carrageenophyte with commercial value, as grown in land-based nursery systems. J. Appl. Phycol. 32:1917–1922.
doi.org/10.1007/s10811-020-02091-7
Umanzor, S., Shin, S., Yarish, C., Augyte, S. & Kim, J. K. 2020b. Exploratory evaluation of the effects of Kelpak ® seaweed extract on cultivated kelp Saccharina spp. exposed to sublethal and lethal temperatures. J. World Aquac. Soc. 51:960–969.
doi.org/10.1111/jwas.12687
Van Staden, J., Beckett, R. P. & Rijkenberg, M. J. 1995. Effect of seaweed concentrate on the growth of the seedlings of three species of Eucalyptus. S. Afr. J. Bot. 61:169–172.
doi.org/10.1016/S0254-6299(15)30513-5
Waaland, J. R., Dickson, L. G. & Watson, B. A. 1990. Protoplast isolation and regeneration in the marine red alga Porphyra nereocystis. Planta. 181:522–528.
doi.org/10.1007/BF00193005
Wada, E. 1941. Experimental studies on the life history of Porphyra tenera Kjellm. Bull. Jpn. Soc. Sci. Fish. 10:47–59. Wally, O. S. D., Critchley, A. T., Hiltz, D. & et al 2013. Regulation of phytohormone biosynthesis and accumulation in Arabidopsis following treatment with commercial extract from the marine macroalga Ascophyllum nodosum. J. Plant Growth Regul. 32:324–339.
doi.org/10.1007/s00344-012-9301-9
Wang, S. J. 1985. Cultivation of Porphyra. In : Tseng C. K., Wang S. J., Liu S. J., editors
Seaweed Cultivation
. Shanghai Science and Technical Publishers, Shanghai, 145–150.
Wang, W.-J., Zhu, J.-Y., Xu, P. & et al 2008. Characterization of the life history of Bangia fuscopurpurea (Bangiaceae, Rhodophyta) in connection with its cultivation in China. Aquaculture. 278:101–109.
doi.org/10.1016/j.aquaculture.2008.01.008
Watanabe, S., Matsumoto, M., Hakomori, Y., Takagi, H., Shimada, H. & Sakamoto, A. 2014. The purine metabolite allantoin enhances abiotic stress tolerance through synergistic activation of abscisic acid metabolism. Plant Cell Environ. 37:1022–1036.
doi.org/10.1111/pce.12218
Yoshie, Y., Suzuki, T., Shirai, T. & Hirano, T. 1993. Free amino acids and fatty acids composition in dried nori of various culture locations and prices. Nippon Suisan Gakkaishi. 59:1769–1775.
doi.org/10.2331/suisan.59.1769
|
|












