Abbreviations
ACE
angiotensin converting enzyme
ALT
alanine aminotransferase
AST
aspartate aminotransferase
BDNF
brain derived neurotrophic factor
CK
creatine kinase
GABA
γ-aminobutyric acid
GFST
GABA-enriched fermented sea tangle
HDL
high-density lipoprotein
HGH
human growth hormone
IGF-1
insulin-like growth factor-1
IL
interleukin
LDL
low-density lipoprotein
mTOR
mammalian target of rapamycin
NAC
acetylcysteine
PSGL-1
Pselectin glycoprotein ligand 1
SO
sarcopenic obesity
TFM
total fat mass
TG
triglyceride
TLM
total lean mass
TNF-α
tumor necrosis factor α
TrK8
tyrosine kinase B
INTRODUCTION
The progressive loss of muscle mass and volume, termed sarcopenia, often leads to functional deterioration and subsequent increases in the incidence of frailty, falls and disability, commonly coupled with high levels of adiposity (sarcopenic obesity [SO]). The multi-factorial nature of SO, mediated by numerous mechanisms, including alpha-motor neuron death, altered hormone concentrations, increased inflammation and altered nutritional status (Jones et al. 2009), necessitates comprehensive and efficacious interventions through exercise, nutrition, pharmacology, or otherwise.
Sea tangle is used frequently and widely across China, Japan, and Korea as a seasoning or dietary ingredient. Sea tangle consists of protein, amino acids, minerals, polyphenols, and insoluble cellulose rich in dietary fibre (Cha et al. 2011a). Once subjected to a highly specialized lactic acid fermentation process, using Lactobacillus brevis BJ20, glutamic acid within the sea tangle is bio-converted into γ-aminobutyric acid (GABA). GABA is a non-protein amino acid specifically functioning as an inhibitory neurotransmitter in the central nervous system. GABA has been reported to induce hypotensive effects and provide diuretic and tranquillizer effects (Inoue et al. 2003). Furthermore, supplementation of GABA-enriched fermented sea tangle (GFST) has been linked to liver protection effects (Lee et al. 2010b, Cha et al. 2011b) and improvements in antioxidant levels (Lee et al. 2010a), obesity status and stress management (Okai et al. 1993, You et al. 2009). GABA supplementation has also been associated with an elevation in the levels of human growth hormone (HGH) in healthy (Cavagnini et al. 1980b) and trained men, and through this, may lead to further effects on substrate metabolism, including lipolysis and skeletal muscle protein accretion (Turner et al. 1988, De Palo et al. 2006). An angiotensin-converting enzyme (ACE) inhibiting effect has also been demonstrated following GABA-enriched food ingestion, which could contribute to enhanced skeletal muscle function (Witham et al. 2008).
Loss of neuromuscular integrity is closely associated with sarcopenia (Cruz-Jentoft et al. 2010). Brain derived neurotrophic factor (BDNF) is the most prevalent neurotrophic factor in the central nervous system, and one of a number of proteins responsible for growth, differentiation and survival of neurons. Alterations in BDNF expression and signaling, linked to neurodegeneration, are typically in reference to “brain derived” rather than peripheral sources, however, BDNF is present in blood platelets (Fujimura et al. 2002), muscle cells, vascular beds, macrophages, and leucocytes (Gielen et al. 2003, Cattaneo et al. 2010), where it may be synthesized and released. Reduced serum BDNF has been soundly implicated in the pathogenesis of neurodegenerative diseases (Ventriglia et al. 2013) including dementia (Lee et al. 2009), Alzheimer’s disease (Laske et al. 2007), and Parkinson’s disease (Scalzo et al. 2010). Furthermore, it has been put forward that skeletal muscle-derived BDNF has a protective function on innervating motor neurons, potentiates neuromuscular transmissions and, of particular interest, stimulates protein synthesis (Oppenheim et al. 1992, Zhang and Poo 2002). Thus, on the basis of previous literature, we hypothesized that the consumption of GFST would increase the levels of blood serum biomarkers associated with muscle building and body fat status, while investigating neuromuscular survival via increased BDNF.
MATERIALS AND METHODS
Participants
Twenty-one middle-aged women (53-63 y) participated in this randomized, double-blind, placebo controlled study (Table 1). Subjects whom suffered with hypertension, chronic degenerative disease (arthritis), diabetes, obesity, and cardiovascular disease were excluded from the study. Adverse reaction to supplementation prior to the experimental period or inability to participate due to particular medication resulted in exclusion. All subjects were physically active taking part in moderate aerobic exercise (1-2 times per week). All participants completed a written informed consent to participate in this study, which was approved by Kyungsung University Ethics Committee (KSU-14-05-002).
Preparation of GFST by Lactobacillus brevis BJ20 and placebo
GFST was prepared by a previously adopted, but modified, procedure (Park et al. 2016). Sea tangle was added to water at a ratio of 1 : 15 (w/v) with the addition of yeast extract and glucose, based on the amount of sea tangle added to the mixture. After autoclaving at 121°C for 30 min, a sea tangle solution was obtained and the Lactobacillus brevis BJ20 (accession No. KCTC 11377BP) culture broth was added to the solution at a concentration of 1.2% (v/v) (Table 2), which was mixed and incubated at 37°C for 2 days (Fig. 1). A soft capsule was made for the clinical trial by mixing 250 mg fermented sea tangle, 61 mg lactose, 32 mg cellulose, 5 mg HPC, 30 mg SiO2, and 7 mg magnesium stearate. A placebo was made by mixing 311 mg lactose, 32 mg cellulose, 5 mg HPC, 30 mg SiO2, and 7 mg magnesium stearate.
During the preparation process, the GABA content had been controlled to range between 40-60 mg within 1,000 mg of GFST. Based on high performance liquid chromatography analysis, it was confirmed that the mean content of GABA was 54.5 ± 0.071 mg g-1 in the GFST used in this study.
General protocol
Over the experimental period, participants ingested ei-ther 1,000 mg of GFST (n = 10) or a sucrose placebo (CON) (n = 11) everyday, for 8 weeks. Subjects were asked to abstain from any regular exercise. Fasting venous blood samples were taken before and after the 8 weeks period.
Body composition
Measurements of total lean mass (TLM) and total fat mass (TFM) were carried out at the Medical Centre for Health Promotion (Busan, Korea) using the DEXA (Dual Energy X-ray Absorptiometry; Discovery QDR Hologic, Bedford, MA, USA). Subjects were fitted with robes and had their measurements taken by professionally trained assistants, taking 15 min for the entire body.
Biochemical analysis on blood
Fasting venous blood samples were taken before and after the 8 weeks period. Collected blood samples were clotted for 30 min centrifugation for 15 min at 1,000 ×g separated into serum and stored at -80°C and analyzed by immunoassay.
Serum substrates and inflammation factors
Glucose substrate is an indispensable energy supplier which supports cellular function. Glucose degradation occurs in glycolysis. The hexokinase method, based on the work of Schmidt, Peterson and Young, is a recognized reference method according to the manufacturer’s procedures (COBAS; Roche Diagnostics, Indianapolis, IN, USA).
Concentrations in serum of triglyceride (TG), high-density lipoprotein (HDL) cholesterol, and low-density lipoprotein (LDL) cholesterol were enzymatically measured with commercial kits (COBAS; Roche Diagnostics).
Tumor necrosis factor α (TNF-α) Immunoassay to measure TNF-α in serum, was performed by using a Quantikine HS ELISA kit provided from R&D Systems, Inc. (Minneapolis, MN, USA). It is insensitive to the addition of recombinant forms of either of the two types of soluble receptors. It is probable that this measurement detects the total amount of TNF-α in serum, i.e., the total amount of free TNF-α plus the amount of TNF-α bound to soluble receptors. Optical density was determined at a wavelength of 490 nm.
Immunoassay to measure interleukin (IL)-6 in serum was performed by using a Quantikine HS ELISA kit provided from R&D Systems, Inc. Soluble forms of IL-6 R alpha are generated by both alternative splicing and proteolytic cleavage. In a mechanism known as trans-signaling, complexes of soluble IL-6 and IL-6 R alpha elicit responses from gp130-expressing cells that lack cell surface IL-6 R alpha.
Growth-related factors
For measuring BDNF concentrations, a commercially available kit from R&D Systems, Inc. was used. Diluted serum was used to determine the optical density of each sample well within 30 min at 450 nm.
ACE mediates the cleavage of the synthetic substrate (FAPGG; N-[3-(2-furyl)acryloyl]-L-phenylalanyl-L-glycyl- L-glycine) into an amino acid derivate and a dipeptide. The kinetic of this cleavage reaction is measured by recording the decrease in absorbance at 340 nm using a commercially available kit from BÜHLMANN Laboratories (Basel, Switzerland).
HGH concentration was determined using, a commercially available kit, Immulite 2000 from Siemens AG (Muenchen, Germany). A growth hormone releasing hormone + arginine stimulation test calibrated with both IS 80/505 and IS 98/574 (GRH Growth hormone-Recombinant 98/574-kit).
Insulin-like growth factor-1 (IGF-1) concentrations were measured using a solid-phase, enzyme-labeled chemiluminescent immunometric assay on the Immulite 2000 automated immunoanalyzer (Siemens AG).
Strength measurements
Isokinetic strength was measured using a Biodex machine (Biodex System 4 Pro; Biodex, Shirley, NY, USA). With subjects seated, the dynamometer was adjusted to match the rotational axis of the worked extensors and flexors. Participants were encouraged to remain seated, not moving other body parts except for the pelvis, with additional external forces applied (straps) to prevent movement of the femoral site and chest. Attachments were used to accommodate for leg length ranges of motion. Isokinetic knee flexion and extension were measured in the anatomical position, through 90°-0° and again from 0°-90° whilst being adjusted for hyperextension. Load speeds of 60° s-1 were performed on the lower limbs to assess muscle strength. All strength measurements were recorded within 7 days of starting the experimental period and 3 days of completion.
Indicators of fatigue and damage on liver and muscle
Aspartate aminotransferase (AST, or glutamate oxalacetate transaminase) and alanine aminotransferase (ALT, or glutamate pyruvate transaminase) both belong to the transaminases, which catalyze the interconversion of amino acids and α-keto acids by transfer of amino groups. A standardized procedure provided by the manufacturer (COBAS; Roche Diagnostics) was used for AST and ALT determination. ALT determination included optimization of substrate concentrations, employment of Tris buffers, preincubation of a combined buffer and serum to allow side reactions with NADH, substrate start and pyridoxal phosphate activation. AST measurement, included optimization of substrate concentrations, employment of Tris buffers, preincubation of a combined buffer and serum to allow side reactions with NADH to occur, substrate start, and optional pyridoxal phosphate activation.
Creatine kinase (CK) is rapidly inactivated by oxidation of the sulfhydryl groups in the active center. The enzyme can be reactivated by the addition of acetylcysteine (NAC). Interference by adenylate kinase is prevented by the addition of didenosine pentaphosphate and AMP. Standardized methods for the determination of CK were used: “reverse reaction” and activation by NAC according to the manufacturer’s method (COBAS; Roche Diagnostics).
Statistics
All statistical analysis was conducted with Statistical Package for Social Sciences (SPSS Windows ver. 18.0; SPSS Inc., Chicago, IL, USA). Descriptive statistics (mean and standard deviation) were used to report on the measurements of each variable. A student paired t test and ANCOVA analysis were conducted to identify within and between group differences. Statistical significance was set at α = 0.05.
RESULTS
Lipid and inflammation
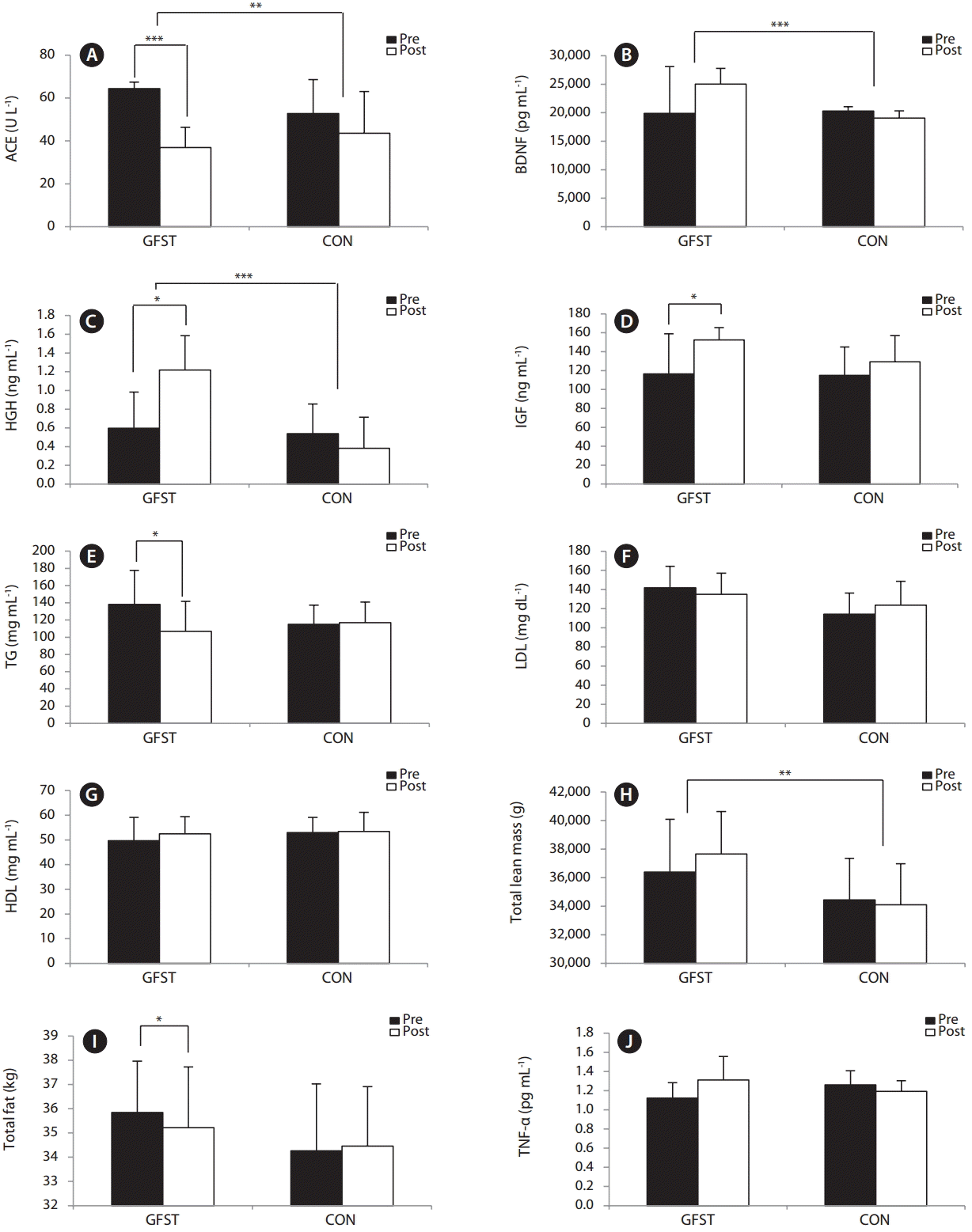
There was no significant effect of GFST on blood glucose. We found a reduction in TG levels in the GFST group (p < 0.05) (Fig. 2E). There were, however, no significant changes in LDL or HDL levels observed, though a notable increase was seen in the latter (Fig. 2G). There were no significant changes in IL-6 or TNF-α levels (Fig. 2J).
BDNF and growth-related factors
No significant within-group effect was observed in BDNF serum levels, but a between-group difference (Fig. 2B) was observed (p < 0.001). A significant reduction was observed in the GFST (p < 0.001) group with respect to ACE concentrations, and between groups (p < 0.01) (Fig. 2A). There was, however, a significant effect of supplementation on HGH levels (p < 0.05), also significantly different between groups (p < 0.001) (Fig. 2C). IGF-1 increased significantly in the GFST experimental group (p < 0.05) (Fig. 2D).
Muscle strength
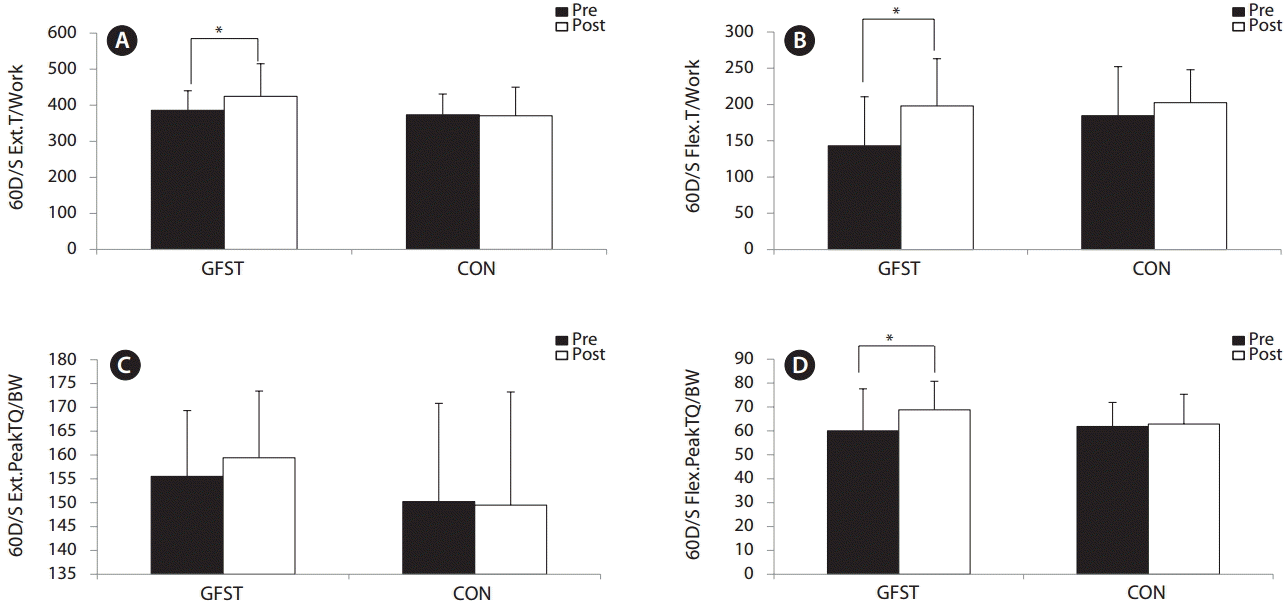
Muscle strength of the lower limb was shown to significantly improve in the GFST group for total work, knee extension and flexion at 60° s-1 (p < 0.05), and peak torque normalized to body weight of knee flexion at 60° s-1 (p < 0.05) (Fig. 3A, B & D, respectively). Peak torque normalized to body weight of knee extension also tended toward increasing but didn’t reach significance.
DISCUSSION
Sea tangle is an ingredient rich in vitamins, minerals, essential fatty acids, carotenoids and bioactive compounds such as alginates and fucoidans. Supplementation of sea tangle has been associated with numerous health benefits, including anti-hyperlipidemia and anti- atherosclerosis effects (Lee et al. 2011), hypotensive (Chiu and Fung 1997) and antioxidant activities (Han et al. 2002).
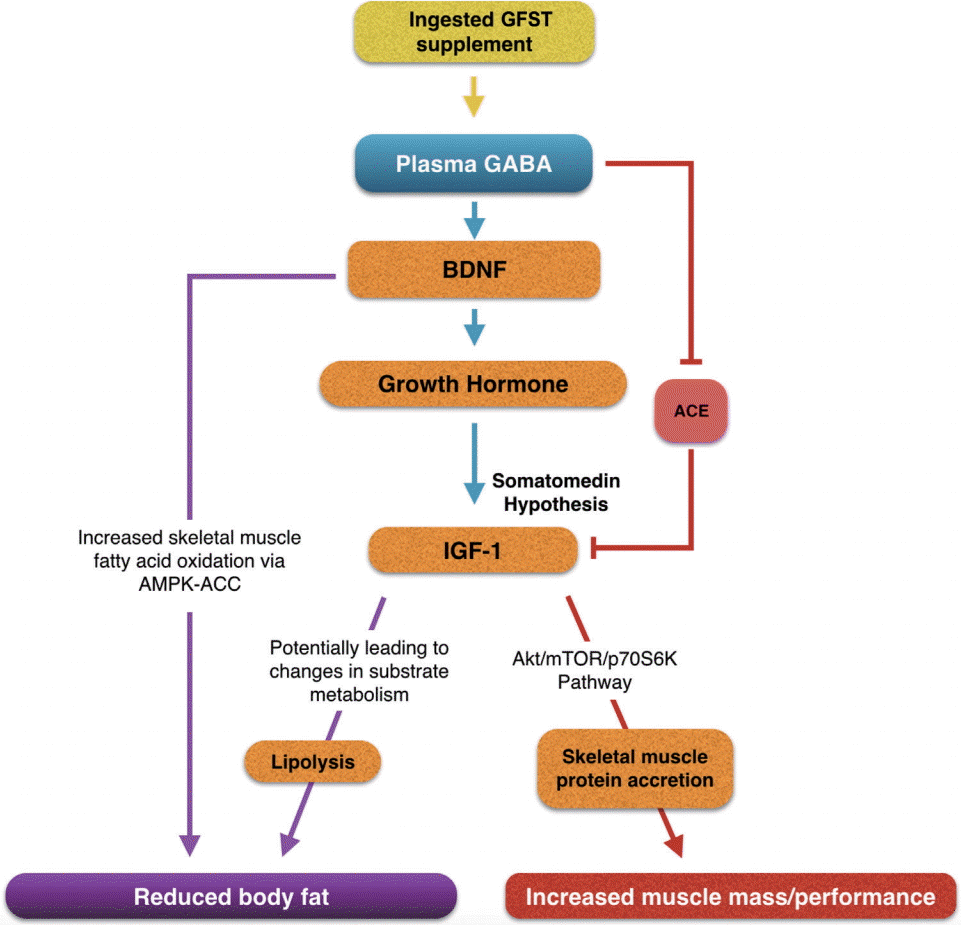
Sea tangle was fermented to enrich the GABA content and evaluate the potential health benefits on SO. The prevailing research has focused on the acute changes induced by oral GABA administration with few studies on functional food intake, and to our knowledge, few reports have rigorously tested the effects of GABA rich functional foods on other biomarkers associated with neuromuscular function, body composition, blood glucose, and cholesterol status. The findings of the present study show an increase in growth factors and improvements in total body fat and lean muscle status, following GFST supplementation, via multi-mechanistic pathways (Fig. 3). Of notable interest was the increased BDNF in the experimental group along with muscle growth factors, ACE, HGH, and IGF-1.
The role of neurotrophic growth factors in neurodegenerative conditions has been the predominant focus of research surrounding nervous system health and function. However, BDNF is attracting attention for its association with neuromuscular disorders and sarcopenia. The Akt/protein kinase B activated mammalian target of rapamycin (mTOR) pathway has emerged as a key regulator of skeletal muscle hypertrophy (Lai et al. 2004), and shows a decreased activation (Kimball et al. 2004) and phosphorylation status with ageing (Haddad and Adams 2006). Muscle membrane tyrosine kinase B (TrKB) receptors are thought to play a role in providing a binding site for BDNF to activate the Akt/mTOR/p70S6K signaling pathway leading to protein synthesis (Sakuma and Yamaguchi 2010). In this study we demonstrated that GFST increased serum BDNF levels when baseline values were not significantly different. This indicates GFST as a viable functional food which may create an environment conducive for muscle protein synthesis. In addition, research has shown the fat oxidation AMPK-ACC signaling pathway to be enhanced as a result of BDNF induced phosphorylation in skeletal muscle (Pedersen 2011) potentially via acting in an autocrine and / or paracrine manner within skeletal muscle (Pedersen 2013). BDNF, also being low in obesity, implies a role for BDNF in the regulation of central and peripheral metabolism, potentially linked to the reduction in TFM observed in the present study. Taken together, BDNF may be identified as a myokine that influences myogenesis, muscle regeneration, and metabolism (Pedersen 2013). More studies are warranted to establish a role for GABA-related BDNF expression in SO, investigating the longitudinal effects of GABA supplementation on BDNF, and whether these translate to improvements in body composition and physical function in the ageing population.
HGH plays a key role in the maintenance and growth of skeletal muscle, and impacts the metabolism of lipids, carbohydrate, and protein in nearly all body tissue. Particular functions of HGH include the stimulation of protein anabolism (i.e., increased DNA, RNA, and protein synthesis), enhanced utilization of fats by stimulating triglyceride breakdown and oxidation of adipocytes and antagonism of insulin action (a key regulator of carbohydrate metabolism). HGH is purported to gradually decline with age at a rate of ~1% per year after the age of 30 (Hermann and Berger 2001), and can be as much as 20-fold less in older than in younger adults (Ryall et al. 2008). The age-related decline in HGH, coupled with lower IGF-1 levels, the key mediator of HGH action, have also been associated with sarcopenia and detrimental changes in body composition (Ferrucci et al. 2002). The results of the current study demonstrated a significant increase in HGH as a result of GFST supplementation. Some of the early GABA research on oral supplementation showed significant elevation in plasma HGH levels in healthy men (Cavagnini et al. 1980a). It is important to note that the majority of research investigates the acute changes in HGH levels (Cavagnini et al. 1980a) whereas, the current study presents a case for long term increases in resting HGH with continual supplementation of GABA; a within and between groups effect of GFST was observed after 8 weeks (Fig. 2C). A HGH induced generation of IGF-1, in accordance with the somatomedin hypothesis (Le Roith et al. 2001), may be responsible for the significant increase in IGF-1 (Fig. 2D) (Laron 1996), and thus, an ensuing anabolic or lipolytic response (De Palo et al. 2006) demonstrated by an increase in TLM (Fig. 2H) and decrease in TFM (Fig. 2I) following GFST supplementation. Furthermore, inferring a stimulatory effect of GABA on HGH levels, the tendency toward a reduction in LDL and increase in HDL concentrations shown in the GFST group (Fig. 1G), help support the notion that following HGH release IGF-1 has a modulating role on insulin sensitivity (Moses et al. 1996) and HDL levels further downstream (Succurro et al. 2010). The relative contributions of these IGF-1 pathways require further investigation, due to the ubiquitous nature of IGF-1, IGF-1, and HGH independent growth stimulating effects (Laron 2001), and / or the possibility of GABA directly stimulating hypothalamic responses (i.e., ghrelin, growth hormone-releasing hormone, and somatostatin) (Powers 2012).
Our results may give an important contribution to the question of whether ACE inhibitor use can aid in sarcopenia treatment. The renin-angiotensin system is known to be activated in many catabolic conditions, in which ACE is responsible for converting angiotensin I to angiotensin II, in a process that regulates blood pressure and the body’s fluid and sodium balance. Angiotensin II infusion resulted in cachexia and decreased circulating IGF-1. The weight loss was associated with skeletal muscle catabolism, in the presence of unchanged fat tissue, possibly via its inhibitory effect on the autocrine IGF-1 system (Brink et al. 2001), and more specifically, via the Akt/mTOR/p706SK pathway (Song et al. 2005). Thus, the inhibitory effect of GABA on ACE demonstrated in the present study, and previously evidenced in the literature (Lin and Omori 2002), points toward a major mechanistic pathway to target in catabolic conditions such as sarcopenia. The extent to which ACE inhibition contributed to the increase in IGF-1 or TLM, in the current study, is unknown, although it may act synergistically with other mechanisms (Fig. 4). Future investigation ought to be carried out on the effects of ACE inhibitors on muscle-specific IGF-I expression in healthy, ageing subjects. Of particular interest to the application of functional foods as lipid status improving agents, was the reduction in TFM (p < 0.05), increased HDL concentration and significant decrease in TG levels (p < 0.05) following GFST supplementation. Brown seaweeds are known to be rich in polysaccharides such as fucoidan, laminarin, and alginate. Fucoidan of brown seaweed has been implicated in the reduction of blood lipids (Li et al. 2008), decrease in total cholesterol, triglycerides, and LDL concentration, and increase in HDL concentration (Li et al. 2001). In addition, reduced cholesterol and triglycerides has been reported in patients with hyperlipidaemia following fucoidan ingestion (Wang and Bi 1994). Up-regulation of lipid metabolic enzymes (Huang et al. 2010) is one proposed mechanism by which fucoidan may help to improve lipid profiles. Obesity can be expressed, partly, as a state of chronic low grade inflammation in adipose tissue, caused by adipocyte hypertrophy and hyperplasia, marked by an increase in macrophage content and activity in the adipose tissue, inducing a release of cytokines and pro-inflammatory signals. Leukocyte trafficking into adipose tissue is mediated by increased endothelial expression of P-selectins and E-selectins (Nishimura et al. 2008), and the regulatory role of P-selectin glycoprotein ligand 1 (PSGL-1) on the adhesive properties of the endothelium and leukocyte. PSGL-1 deficiency was shown to have protective effects against visceral fat inflammation in mice, by reducing IL-1 receptor-mediated adhesive properties of the endothelium (Russo et al. 2010). Furthermore, antibody blockade of P- and E-selectin reduced leukocyte rolling and firm attachment in obese visceral adipose tissue (Nishimura et al. 2008). Fucoidan acts as a “selectin blocker” (Kylin 1913) and therefore may reduce selectin-dependant leukocyte rolling on endothelial cells, subsequently leading to a reduced inflammatory state. It is known that leukocyte and neutrophils correlate positively with increased visceral fat in obese individuals (Ryder et al. 2014), thus targeting the adhesive properties of endothelial cells may prove an efficacious treatment for inflammatory disease states.
Another property of sea tangle is alginate, a dietary fibre extracted from the cell walls of brown seaweed. Studies have reported a reduction in body fat percentage(Georg et al. 2012), acute modulations in human-appetite sensation (Paxman et al. 2008) and decreased gastric emptying rate attributed to alginate ingestion. Inhibition of digestive enzymes in vitro (namely, pancreatic lipase) (Wilcox et al. 2014) and altered intestinal content viscosity are proposed key factors mediating the uptake of triacylglycerol and promoting the control of energy intake (Kristensen and Jensen 2011), respectively. These effects are still inconclusive (Odunsi et al. 2010) and warrant further investigation. Special attention should be given to the effectiveness of alginates extracted from different types of seaweed, the dose-dependency of alginate usage (the more alginate the less enzyme activity) and the palatability of potential supplements.
To our knowledge, no previous studies have investigated the effects of GABA-enriched functional foods on performance outcomes, particularly strength. Our findings of significantly improved isokinetic strength (Fig. 3A, B & D) in the GFST group help support the stance that functional foods, such as GFST, can significantly enhance growth factor levels, such as HGH and IGF-1, resulting in functional improvements in performance. The inclusion of functional foods as part of a holistic intervention seems an efficacious, noninvasive and practical approach in tackling SO and improving the ability to carry out active daily living tasks with age. It can also be added that no adverse effects to the liver were reported following GFST supplementation (Appendix 1). In this study, we demonstrated that GFST significantly decreased TFM and TG, as well as significantly increasing serum BDNF, ACE, HGH, and IGF-1 levels.
In conclusion, the use of functional foods, particularly GFST, to increase the levels of circulating GABA may stimulate the release of muscle-related growth factors including serum BDNF and / or affect substrate metabolism, providing a safe, protective intervention for age-related conditions such as SO. However, further studies concerning GFST as a functional food and its potential use in conditions such as SO should be addressed.